研究背景
胎生是脊椎動物中廣泛存在的一種生殖策略,其特征為受精卵在母體內(nèi)發(fā)育,直至胚胎發(fā)育成熟后才被產(chǎn)出。與這一普遍模式形成鮮明對比的是,海馬科物種演化出了極為特殊的“雄性懷孕”現(xiàn)象,該獨特的生命史策略為研究動物生殖系統(tǒng)的演化提供了重要窗口:功能性相似的生殖方式究竟源于趨同的分子機制,抑或是通過同源通路或全新的細(xì)胞調(diào)控路徑實現(xiàn)?盡管已有比較基因組學(xué)研究初步揭示了該生殖方式轉(zhuǎn)變的遺傳背景,然而相關(guān)基因在特定細(xì)胞類型中的表達模式及其演化軌跡仍不清楚。
在結(jié)構(gòu)上,海馬育兒袋與有袋類動物的育兒袋具有一定相似性,并在功能上融合了羊膜動物子宮與胎盤的特點。在雄性懷孕過程中,其育兒袋內(nèi)層組織發(fā)生顯著肥大與血管化,形成一種被稱為“偽胎盤”的結(jié)構(gòu),可執(zhí)行氣體交換與營養(yǎng)物質(zhì)傳遞等典型的胎盤功能。此外,海馬在進化過程中丟失了?foxp3?基因——該基因在哺乳動物中對調(diào)節(jié)性T細(xì)胞(Treg)的發(fā)育與功能維持具有核心作用,這一遺傳缺失也引發(fā)了對其在懷孕過程中免疫耐受機制的特殊適應(yīng)性的探討。
研究內(nèi)容和結(jié)果
對海馬育兒袋7個發(fā)育階段進行單細(xì)胞轉(zhuǎn)錄組測序,鑒定出14個細(xì)胞簇,分為四大主要細(xì)胞類型:上皮細(xì)胞(EPCs,9個簇)、成纖維細(xì)胞(FBs,2個簇)、免疫細(xì)胞(ICs,2個簇)和內(nèi)皮細(xì)胞(ENCs,1個簇)。細(xì)胞的空間分布對細(xì)胞間相互作用及其功能維持至關(guān)重要。
對妊娠早期胎盤囊進行空間轉(zhuǎn)錄組測序(測序平臺BMKMANU?S1000)發(fā)現(xiàn),EPCs在胎盤囊的三層結(jié)構(gòu)中均呈現(xiàn)高豐度。其中,EPCs-II(高表達C型凝集素)在外層富集,可能暗示其在早期聚集外部吸附物或抑制表皮細(xì)菌方面發(fā)揮作用。相比之下,EPCs-III(tfa+)和EPCs-IV(gata1+)在內(nèi)層富集,與“鐵穩(wěn)態(tài)”、“細(xì)胞遷移”及“血管發(fā)育”相關(guān),其在雄性妊娠胎盤形成過程中可能參與侵襲和血管化過程。作為細(xì)胞外基質(zhì)(ECM)的主要來源,F(xiàn)Bs存在于中間層,富集“膠原蛋白生成”,可能促進胎盤囊結(jié)構(gòu)形成和組織重塑。ICs則分散于三層結(jié)構(gòu)中,調(diào)控免疫穩(wěn)態(tài)。
基于scRNA-seq,scATAC-seq,空間轉(zhuǎn)錄組數(shù)據(jù)聯(lián)合分析,作者發(fā)現(xiàn)了具有干細(xì)胞潛能的”育兒袋上皮祖細(xì)胞(BEPCs)”。研究顯示,該類細(xì)胞在發(fā)育過程中與膠原蛋白基因呈現(xiàn)協(xié)同表達,并受到雄激素信號的強烈驅(qū)動。研究進一步證實,外源性雄激素處理可誘導(dǎo)雌性海馬形成育兒袋結(jié)構(gòu)。由此,雄激素受體及其調(diào)控的育兒袋上皮祖細(xì)胞作為觸發(fā)育兒袋器官生成的關(guān)鍵起始因子。
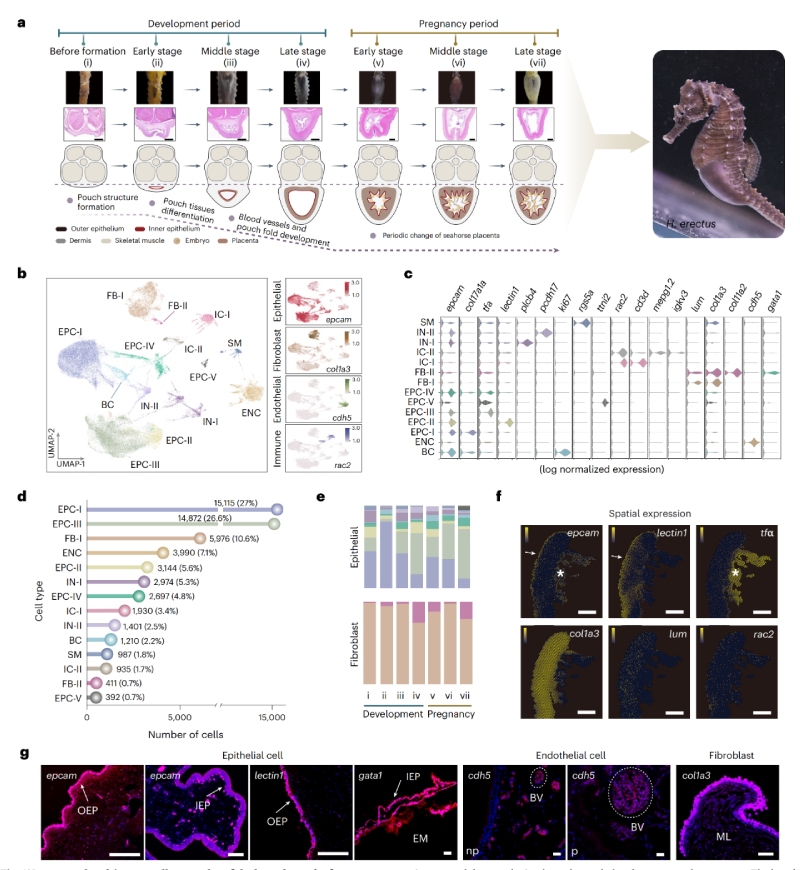
圖1?細(xì)胞圖譜構(gòu)建
在雄性妊娠期間,海馬卵囊內(nèi)層會發(fā)生顯著的形態(tài)學(xué)改變并呈現(xiàn)肥大,形成類似胎盤的組織結(jié)構(gòu),在細(xì)胞譜系轉(zhuǎn)變過程中,多個海馬特異性基因(如pastn1、sp-chia)在偽胎盤形成中起關(guān)鍵作用。作者發(fā)現(xiàn)EPC-I(muc15)、EPC-I(cxcl14)和EPC-IV在人類中與絨毛外滋養(yǎng)層細(xì)胞(EVTs)及絨毛滋養(yǎng)層細(xì)胞(VCTs)具有高度相似的基因表達譜,海馬雄性妊娠和哺乳動物常規(guī)雌性妊娠的胎盤形成過程,可能具有趨同的細(xì)胞機制和調(diào)控基礎(chǔ)。海馬缺乏foxp3基因(哺乳動物中調(diào)控Treg細(xì)胞的關(guān)鍵基因),但可能通過其它免疫細(xì)胞(如cd4+il2rb+?Tregs?和巨噬細(xì)胞)維持對胚胎的免疫耐受。
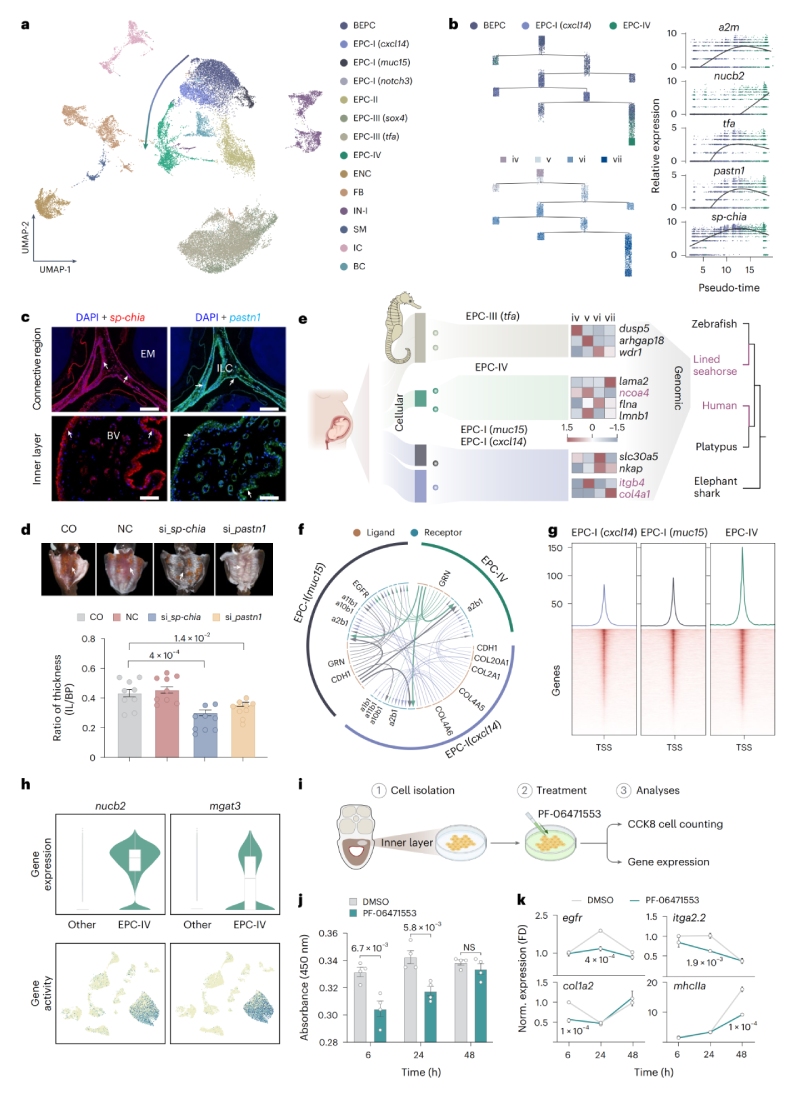
圖2?海馬假胎盤形成與子宮重塑的單細(xì)胞轉(zhuǎn)錄組分析圖譜
通過跨物種比較基因組學(xué)與單細(xì)胞多組學(xué)分析發(fā)現(xiàn),盡管存在物種特異性差異,海馬假胎盤中的多數(shù)細(xì)胞類型在轉(zhuǎn)錄組特征上高度近似于人類的滋養(yǎng)層細(xì)胞——后者在調(diào)控胎兒生長及母體妊娠適應(yīng)中發(fā)揮關(guān)鍵作用。其中,EPCs-II型細(xì)胞兼具表皮分化特征與凝集素介導(dǎo)的黏附功能,其可能是卵囊演化起源的重要線索。
進一步研究表明,海馬育兒袋與哺乳動物子宮在細(xì)胞和遺傳層面具有顯著同源性。海馬與哺乳動物生殖系統(tǒng)所呈現(xiàn)的趨同進化現(xiàn)象,可能源于特定細(xì)胞類型通過趨同演化形成相似的轉(zhuǎn)錄特征,進而實現(xiàn)類似功能,最終推動胎生機制的形成。
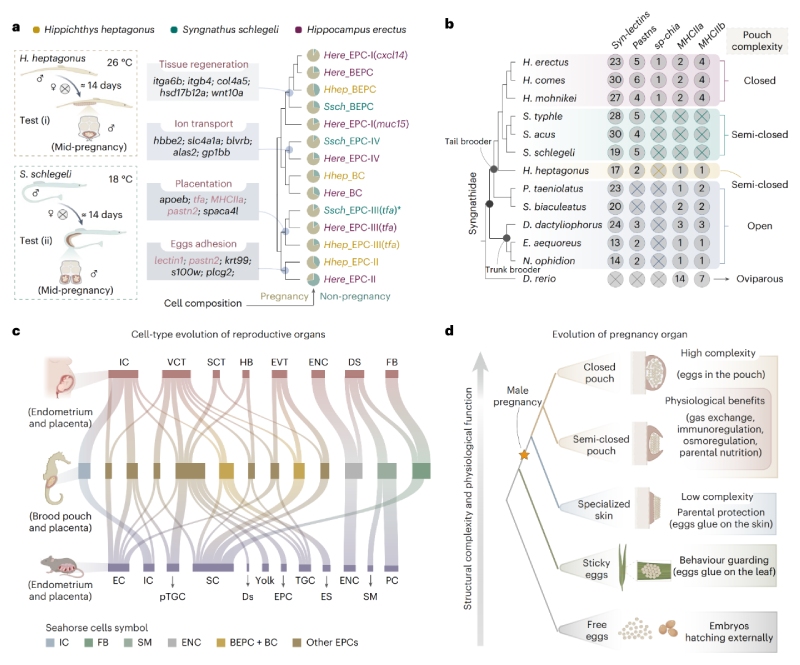
圖3?細(xì)胞類型進化分析
研究總結(jié)
該研究通過細(xì)胞分子與發(fā)育生物學(xué)的多維度分析,深入解析其卵囊發(fā)育與妊娠過程中的細(xì)胞遺傳動態(tài)。研究發(fā)現(xiàn),卵囊形成的關(guān)鍵在于一種具有干細(xì)胞潛能的“育兒袋上皮祖細(xì)胞”群體。體內(nèi)實驗證實,雄激素在卵囊形成中起主導(dǎo)作用。通過對其它動物的對比研究,作者揭示了海馬雄性親代撫育的早期進化機制,以及其細(xì)胞特征,存在與哺乳動物胎生動物相似的生命功能的趨同演化。
]]>環(huán)境配置
1. conda安裝
wget?https://repo.anaconda.com/miniconda/Miniconda3-py39_4.12.0-Linux-x86_64.sh
下載完成之后運行(按提示安裝)
sh Miniconda3-py39_4.12.0-Linux-x86_64.sh
安裝完成后執(zhí)行以下命令
source ~/.bashrc
2. 環(huán)境配置
conda create -n (環(huán)境名) python=3.9
激活環(huán)境:
conda activate (環(huán)境名)
添加鏡像源
conda config –add channels?https://mirrors.bfsu.edu.cn/anaconda/cloud/bioconda/
conda config –add channels?https://mirrors.bfsu.edu.cn/anaconda/cloud/conda-forge/
conda config –add channels?https://mirrors.bfsu.edu.cn/anaconda/pkgs/free/
conda config –add channels?https://mirrors.bfsu.edu.cn/anaconda/pkgs/main/
查看鏡像源
conda config –show channels
3. 安裝python模塊
conda install numpy=1.23.4
conda install opencv=4.7.0
conda install pandas=2.2.2
conda install matplotlib=3.8.4
conda install tqdm=4.66.4
conda install seaborn=0.12.2
conda install tifffile
conda install scanpy=1.9.8
4. 軟件安裝
conda install samtools=1.18 sambamba=1.0.1 bedtools=2.31.1
conda install macs2=2.2.7.1
conda install deeptools=3.5.5
conda install ucsc-bedgraphtobigwig=472
conda install bwa=0.7.17
conda install bioconda::cutadapt=5.0
conda install bioconda::ucsc-bedsort
conda install bioconda::ucsc-bedclip
5. 安裝指定R版本(v4.3)
conda install -c conda-forge r-base=4.3.1
6. 安裝指定R包
conda install -c conda-forge r-signac=1.14.0
conda install -c conda-forge r-seurat=4.3.0
conda install bioconda::bioconductor-rtracklayer=1.62.0
conda install -c conda-forge r-kableextra=1.4.0
conda install -c conda-forge r-optparse=1.7.5
conda install -c conda-forge r-this.path=2.7.0
conda install conda-forge::r-r.utils
conda install conda-forge::r-devtools=2.4.5
options(timeout = 9999999999999)
devtools::install_github(“davidsjoberg/ggsankey”)
install.packages(‘DT’)
參考基因組配置
參考基因組需要使用bwa進行建庫(流程使用bwa比對)
gtf文件中需包含gene_id,gene_name,gene_biotype/gene_type
人和小鼠基因組推薦使用:
https://www.10xgenomics.com/support/software/cell-ranger-arc/latest/release-notes/reference-release-notes
refdata-cellranger-arc-GRCh38-2020-A-2.0.0
refdata-cellranger-arc-mm10-2020-A-2.0.0
其他基因組可自行選擇版本。
配置文件填寫
配置文件可參考config.txt進行填寫,內(nèi)容說明如下:
## fq數(shù)據(jù)路徑,F(xiàn)Q1為read1文件路徑,F(xiàn)Q2為read2文件路徑
FQ1 /path/to/read_1.fq.gz
FQ2 /path/to/read_2.fq.gz
## 芯片解碼文件路徑
FLU /path/to/flu_info.txt
## 組織明場圖片路徑
HE /path/to/HE.tif
#INSIDE 1 #是否扣除組織內(nèi)部空白區(qū)域,默認(rèn)1扣除,選0為保留
## 組織熒光圖片路徑
#FL /path/to/FL.tif #組織熒光圖,和HE圖二選一
#FLC -1 #熒光圖顏色通道,值為0,1或2,選擇-1為自動判斷
## 組織區(qū)域文件路徑
JSON /path/to/roi.json #如不給定,則流程會自動使用上面的圖片進行識別,自動識別建議使用熒光圖
## 參考基因組文件
GenomeVer xxxx #版本信息,出現(xiàn)在報告中
Ref /path/to/ref/genome.fa #基因組fa文件
Gtf /path/to/ref/gene.gtf #基因組gtf文件
SPECIES human #物種文件,人和小鼠分別為human/mouse,其他物種自行給定
## 輸出結(jié)果及輸出文件前綴
OUTDIR /path/to/result/dir/
PREFIX outfile-prefix
## barcode 類型,S3000芯片無特殊情況為V2
BCType V2
## 線程數(shù)
Threads 8
## cs序列類型,無特殊情況為cs12
CSType cs12
軟件運行
在配置好的環(huán)境中,運行命令:
/path/to/BSATAC -c config.txt -s 0
其中 -c 是配置文件,-s 是所選擇運行的步驟,0是全部運行
結(jié)果說明
outdir/
├── 01.fastq2BcUmi # barcode識別目錄
├── 02.LinkBcChip # 芯片barcode對應(yīng)
├── 03.AllheStat # 多級分辨率和組織區(qū)域處理
├── 04.Cutadapt # 接頭過濾
├── 05.Mapping # reads比對結(jié)果
├── 06.Fragment # Fragment分析
├── 07.WebReport # 網(wǎng)頁版報告
├── BSTViewer_project # 收集的結(jié)果目錄
└── prefix # 芯片原始barcode結(jié)果
聯(lián)合分析操作指南
注意說明,當(dāng)前分析需要百創(chuàng)智造的空間RNA和空間ATAC分析流程輸出的結(jié)果!!!
空間RNA和空間ATAC的主要結(jié)果各自存儲在BSTViewer_project文件下。
1. FiJi對齊
1.1:百創(chuàng)智造空間ATAC與空間轉(zhuǎn)錄組聯(lián)合分析之切片對齊概述
https://zhuanlan.zhihu.com/p/1903040885328945604
1.2:百創(chuàng)智造空間ATAC與空間轉(zhuǎn)錄組聯(lián)合分析之切片對齊前圖像生成-01
https://zhuanlan.zhihu.com/p/1898370805919903880
1.3:百創(chuàng)智造空間ATAC與空間轉(zhuǎn)錄組聯(lián)合分析之FIJI中手動對齊切片-02
https://zhuanlan.zhihu.com/p/1903079101796941928
1.4:百創(chuàng)智造空間ATAC與空間轉(zhuǎn)錄組聯(lián)合分析之切片上點的映射-03
https://zhuanlan.zhihu.com/p/1903081341429452952
1.5:百創(chuàng)智造空間ATAC與空間轉(zhuǎn)錄組聯(lián)合分析之Level1向其他level水平的點映射-04
https://zhuanlan.zhihu.com/p/1907854262597321846
其它: Fiji的下載與安裝(windows 10)
https://zhuanlan.zhihu.com/p/18027378723
特殊說明:軟文詳細(xì)講解了對齊的原理,腳本實現(xiàn)以及結(jié)果說明。其中,軟文中涉及的腳本存在?[SpatialATAC/v1.0.2/fragment/ImageJ_FJ] 中。
2. 聯(lián)合分析
2.1 配置文件說明
配置文件可參考config.yaml?[SpatialATAC/v*/fragment/Joint_analysis/v*/]?進行填寫,內(nèi)容說明如下:
ATAC和RNA分析路徑(一級目錄下)
ATAC_analysis path/BSTViewer_project/
RNA_analysis path/BSTViewer_project/
######映射文件(通過Image Fiji獲取)(默認(rèn)讀取L18 L9 L7 L5四個水平的結(jié)果)Fiji path/
輸出路徑 outdir path/analysis
##其它參數(shù) alpha 0.1 ##透明度,范圍是0-1 logfc 0.1
2.2 流程執(zhí)行
perl Joint_main_v1.2.pl -c config.yaml -s 0
其中 -a -b -c 是配置文件,-s 是所選擇運行的步驟,0是全部運行
perl Joint_main_v1.2.pl
Usage:
-a spatial config ##空間轉(zhuǎn)錄組主流程的配置文件
-b atac config ##空間ATAC主流程的配置文件
-c joint config ##聯(lián)合分析的配置文件(見2.1)
-s run step
step number:
0: run all step
1: Joint analysis
2: Joint report
3: total report
2.3 結(jié)果說明
L*_alignment/
├── L*_Joint_marker_gene.csv ##聯(lián)合分析marker基因
├── L*_joint.rds ##rds文件
├── L*_Sankey_plots.pdf ##桑基圖
├── L*_Sankey_plots.png
├── L*_heatmap.pdf ##marker基因熱圖
├── L*_heatmap.png
├── L*_CoveragePlot.pdf ##基因組瀏覽器圖
├── L*_CoveragePlot.png
├── L*_Spatial_ATAC.cluster.pdf ##ATAC聚類圖
├── L*_Spatial_ATAC.cluster.png
├── L*_Spatial_Joint.cluster.pdf ##聯(lián)合分析聚類圖
├── L*_Spatial_Joint.cluster.png
├── L*_Spatial_RNA.cluster.pdf ##RNA聚類圖
└── L*_Spatial_RNA.cluster.png
**特殊說明**
1. L*_joint.rds 中存在3個assay,分別是:ATAC(peak矩陣),Spatial(表達矩陣),RNA(活性矩陣)。
2. 結(jié)果生成2個報告:聯(lián)合分析的報告,以及Spatial、ATAC以及聯(lián)合分析的匯總報告
研究單位:西安交通大學(xué)基礎(chǔ)醫(yī)學(xué)院院長張保軍教授團隊
期刊名:cellular & molecular immunology
影響因子:21.8
樣本類型:8-12周齡小鼠
文章采用技術(shù):空間轉(zhuǎn)錄組測序、單細(xì)胞RNA測序(scRNA-seq)
引言
在對抗感染和腫瘤的免疫戰(zhàn)場上,記憶?CD8+?T?細(xì)胞是守護機體的「長效衛(wèi)士」,?它們能快速識別再次入侵的病原體或癌細(xì)胞,發(fā)動強力攻擊。長久以來,樹突狀細(xì)胞(DC)被認(rèn)為是主導(dǎo)記憶?T?細(xì)胞分化的核心「指揮官」。研究人員通過整合單細(xì)胞RNA測序(scRNA-seq)和空間轉(zhuǎn)錄(BMKMANU?S1000)技術(shù),揭示了單核細(xì)胞才是調(diào)控記憶?CD8+?T?細(xì)胞分化的關(guān)鍵「操盤手」,其奧秘藏在「細(xì)胞接觸」與「分子信號」的雙重機制中。其中BMKMANU?S1000空間轉(zhuǎn)錄組學(xué)平臺大放異彩,成為揭CCR2+單核細(xì)胞促進記憶CD8+?T細(xì)胞分化的關(guān)鍵工具。
研究背景
CD8+?T細(xì)胞在適應(yīng)性免疫中扮演著關(guān)鍵角色,能夠保護機體免受病原體感染和消除惡性細(xì)胞。當(dāng)CD8+?T細(xì)胞識別到抗原后,會迅速增殖并分化為效應(yīng)CD8+?T細(xì)胞和記憶CD8+?T細(xì)胞。效應(yīng)CD8+?T細(xì)胞負(fù)責(zé)即時清除感染,而記憶CD8+?T細(xì)胞則提供長期保護,能夠在再次遇到相同抗原時迅速啟動免疫反應(yīng)。然而,單核細(xì)胞(monocytes)作為重要的髓系免疫細(xì)胞,雖在炎癥遷移中作用明確,但其是否直接參與T細(xì)胞分化尚無定論。
研究策略
動物模型與樣本制備
實驗動物:優(yōu)先選擇8-12周齡的CD45.1+/CD45.2+及OT-I轉(zhuǎn)基因小鼠
樣本制備:脾臟單細(xì)胞懸液通過機械分離和紅細(xì)胞裂解(ACK緩沖液)制備
技術(shù)手段
- 流式細(xì)胞術(shù)分析
細(xì)胞分選:從感染LM-WT的小鼠脾臟中分選出cDCs、moDCs和單核細(xì)胞等抗原呈遞細(xì)胞(APCs)
細(xì)胞刺激與培養(yǎng):將分選出的APCs與OVA肽段體外共孵育后,過繼轉(zhuǎn)移到已接受初始OT-I+?CD8+?T細(xì)胞的WT受體小鼠體內(nèi),或與體外刺激的初始CD8+?T細(xì)胞共培養(yǎng)
表型檢測:通過流式細(xì)胞術(shù)檢測CD8+?T細(xì)胞的分化表型,包括記憶相關(guān)標(biāo)記物(如cKit、Sca1、Bcl6、TCF1和Eomes)的表達
- 空間分辨轉(zhuǎn)錄組學(xué)
樣本處理:對感染第0天和第5天的小鼠脾臟組織進行切片,并進行H&E染色以確定組織形態(tài)學(xué)特征
空間轉(zhuǎn)錄組學(xué)分析:利用BMKMANU?S1000空間轉(zhuǎn)錄組學(xué)技術(shù)對脾臟切片進行分析,揭示不同免疫細(xì)胞亞群在脾臟中的空間分布
數(shù)據(jù)整合:將scRNA-seq數(shù)據(jù)與空間轉(zhuǎn)錄組學(xué)數(shù)據(jù)進行整合,通過Seurat等工具的FindTransferAnchors和TransferData函數(shù),將scRNA-seq中識別的細(xì)胞類型映射到空間轉(zhuǎn)錄組學(xué)數(shù)據(jù)中,實現(xiàn)細(xì)胞類型的空間定位
- 免疫熒光染色
樣本處理:對感染第0天和第5天的小鼠脾臟組織進行切片,并進行免疫熒光染色
抗體選擇:使用特異性抗體標(biāo)記CD8+?T細(xì)胞、單核細(xì)胞(CCR2+)、樹突狀細(xì)胞(CD11c+)等關(guān)鍵細(xì)胞類型
結(jié)果分析:通過顯微鏡觀察并拍攝染色切片,分析不同細(xì)胞類型在脾臟中的共定位關(guān)系
- 生物信息學(xué)分析
差異基因分析:利用Wilcoxon秩和檢驗等統(tǒng)計方法,識別不同細(xì)胞亞群間的差異表達基因
通路富集分析:通過ClusterProfiler等工具對差異表達基因進行通路富集分析,揭示潛在的生物學(xué)過程
偽時間分析:利用Monocle3等工具構(gòu)建細(xì)胞分化軌跡的偽時間軸,分析CD8+?T細(xì)胞亞群在分化過程中的基因表達變化
研究結(jié)果
研究結(jié)果一:通過空間分辨轉(zhuǎn)錄組學(xué)繪制急性感染后脾組織的圖譜
為了研究免疫反應(yīng)過程中不同免疫細(xì)胞的空間分布和CD8+?T細(xì)胞分化的調(diào)節(jié)機制,研究人員通過整合單細(xì)胞RNA測序(scRNA-seq)和BMKMANU?S1000空間轉(zhuǎn)錄組(ST)技術(shù),分析了急性感染模型中脾臟免疫細(xì)胞的空間分布與CD8??T細(xì)胞分化機制。實驗采用OT-I轉(zhuǎn)基因小鼠模型,通過李斯特菌(LM-OVA)感染誘導(dǎo)免疫反應(yīng),關(guān)鍵發(fā)現(xiàn)包括:1)脾臟形成7個功能集群(B細(xì)胞、T細(xì)胞、單核/巨噬細(xì)胞等);2)感染后T/B細(xì)胞區(qū)擴大,APCs(B細(xì)胞、DC、巨噬細(xì)胞)在T細(xì)胞區(qū)外圍聚集;3)空間重組與T細(xì)胞分化(MP/TE亞群)顯著相關(guān)。多組學(xué)整合揭示了免疫微環(huán)境的動態(tài)調(diào)控機制,為理解感染中細(xì)胞互作提供了多組學(xué)視角。
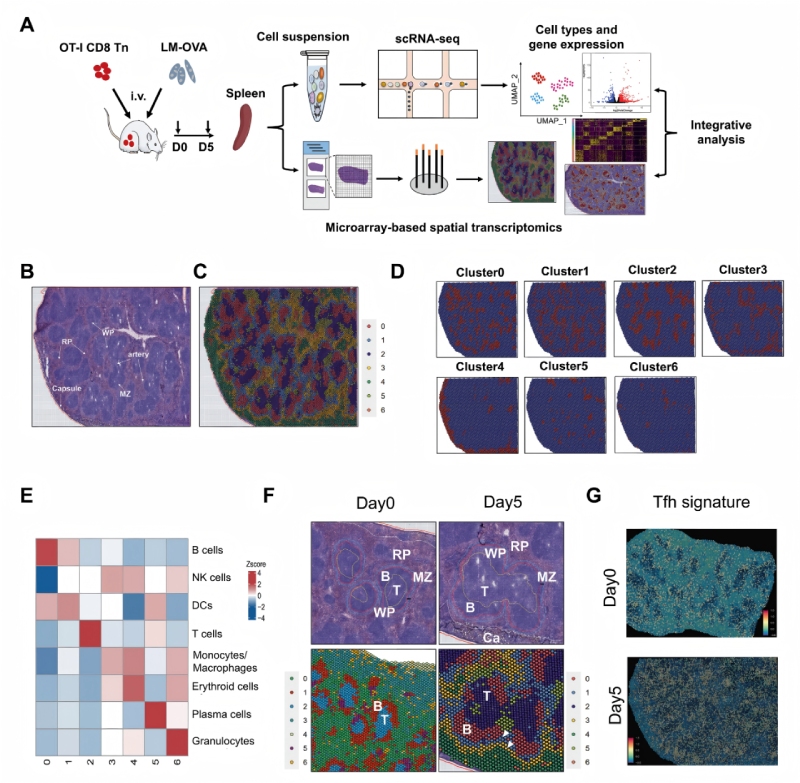
研究結(jié)果二:急性感染期間效應(yīng)和memoryCD8?T細(xì)胞分化軌跡不同
為了探索CD8+?T細(xì)胞分化的空間決定因素,特別是與近端細(xì)胞的相互作用,對于急性感染模型研究人員進行了亞群分析、分化路徑分析和功能驗證,結(jié)果表明在急性感染中,IFN反應(yīng)型CD8+?T細(xì)胞和CD8+MP?細(xì)胞之間分化軌跡的不同意味著,效應(yīng)和記憶CD8+?T細(xì)胞的命運決定于免疫反應(yīng)的初始階段,而IFN應(yīng)答和MP?CD8+?T細(xì)胞分別是效應(yīng)和記憶CD8+?T細(xì)胞分化途徑的初始階段。
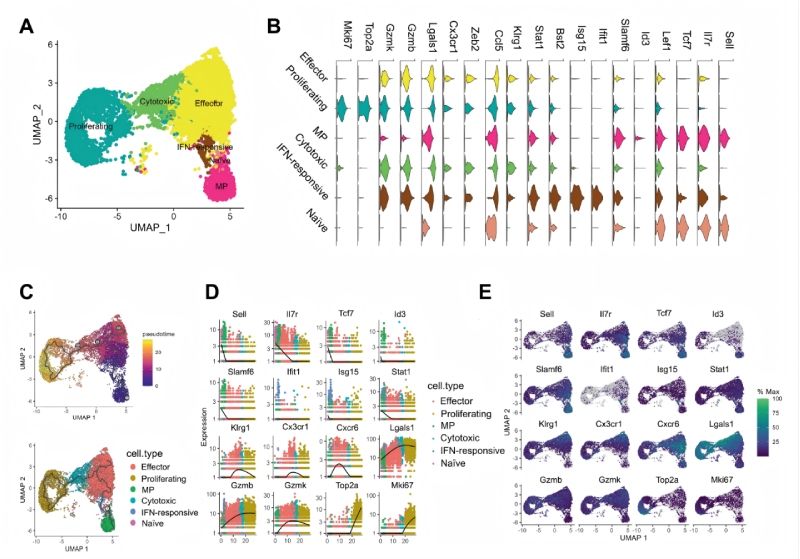
研究結(jié)果三:跨組織區(qū)域細(xì)胞亞群的鑒定和空間圖譜
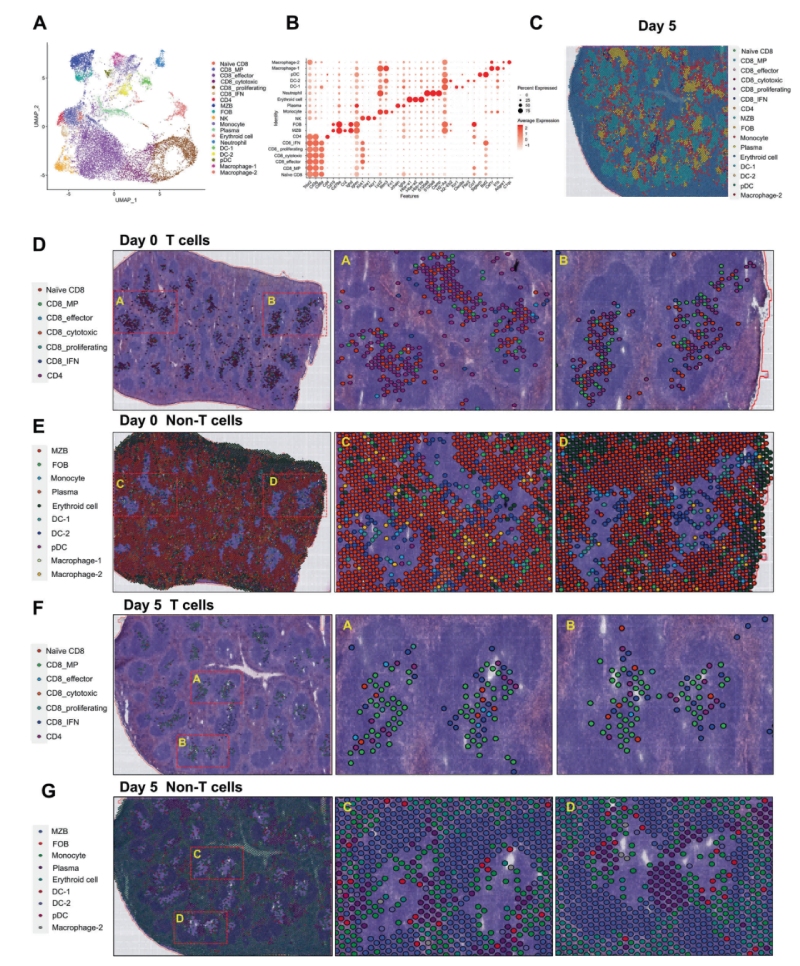
為了研究脾臟免疫細(xì)胞在感染前后的動態(tài)變化,研究人員通過整合單細(xì)胞RNA測序(scRNA-seq)和BMKMANU?S1000空間轉(zhuǎn)錄組(ST)技術(shù),進一步鑒定了供體和受體脾臟中的免疫細(xì)胞群。結(jié)果顯示:①細(xì)胞組成:從1.6萬細(xì)胞中鑒定出19類,包括CD8??T細(xì)胞亞群(如記憶前體和IFN反應(yīng)性細(xì)胞)及髓系細(xì)胞。②動態(tài)變化:感染后第5天,T細(xì)胞區(qū)從以初始CD8+?T細(xì)胞為主轉(zhuǎn)為記憶前體和效應(yīng)細(xì)胞主導(dǎo),且單核細(xì)胞取代DC成為主要浸潤的髓系細(xì)胞。③功能提示:不同的APC和CD8+?T細(xì)胞之間的相互作用在免疫反應(yīng)和CD8+?T細(xì)胞的分化中起著重要作用。
研究結(jié)果四:單核細(xì)胞和CD8+?MP細(xì)胞的抗原依賴性共定位
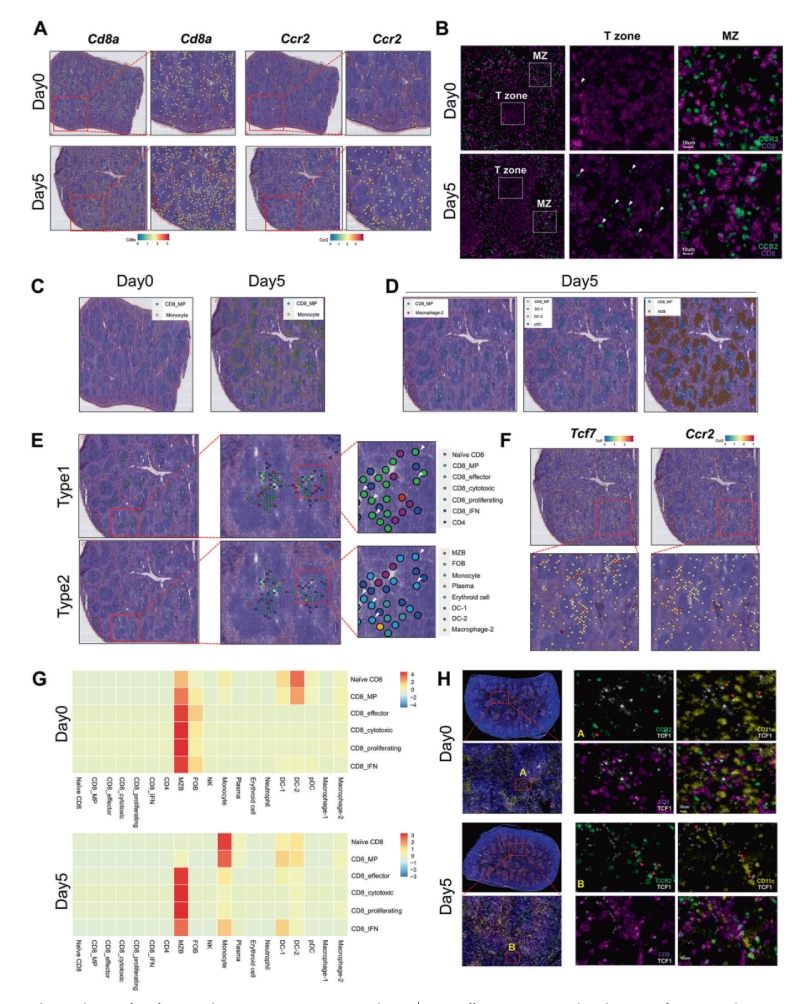
為了探索不同細(xì)胞類型之間潛在的細(xì)胞相互作用,研究人員通過評估特異性免疫細(xì)胞標(biāo)記物的表達、免疫熒光染色及空間轉(zhuǎn)錄組分析,檢查了第0天和第5天脾臟中CD8+?T細(xì)胞和單核細(xì)胞的空間分布。結(jié)果顯示單核細(xì)胞和CD8+?MP細(xì)胞以抗原刺激依賴性的方式在空間上共定位,表明單核細(xì)胞可能對CD8+?MP細(xì)胞的分化有關(guān)鍵影響。
研究總結(jié)
CD8+T?細(xì)胞是適應(yīng)性免疫的重要執(zhí)行者;特別是記憶CD8+?T細(xì)胞對強效和長期保護至關(guān)重要。CD8+?T細(xì)胞分化在轉(zhuǎn)錄水平上的調(diào)控機制已被廣泛研究。然而,人們對感染過程中效應(yīng)和記憶CD8+?T細(xì)胞分化的空間要求仍然知之甚少。研究人員通過整合BMKMANU?S1000空間轉(zhuǎn)錄組(ST)技術(shù)和scRNA-seq技術(shù),發(fā)現(xiàn)了記憶CD8+?T細(xì)胞分化的新機制,該機制涉及單核細(xì)胞和CD8+?MP細(xì)胞之間的解剖學(xué)鄰近性和TGF-β信號傳導(dǎo)。該研究結(jié)果闡述了記憶CD8+?T細(xì)胞命運決定的新機制,并強調(diào)單核細(xì)胞作為關(guān)鍵的APC群體,通過細(xì)胞間接觸依賴的方式在感染過程中促進記憶CD8+?T細(xì)胞的分化。
]]>“療猘犬咬人方,仍殺所咬犬,取腦敷之,后不復(fù)發(fā)”—東晉?葛洪《肘后備急方》
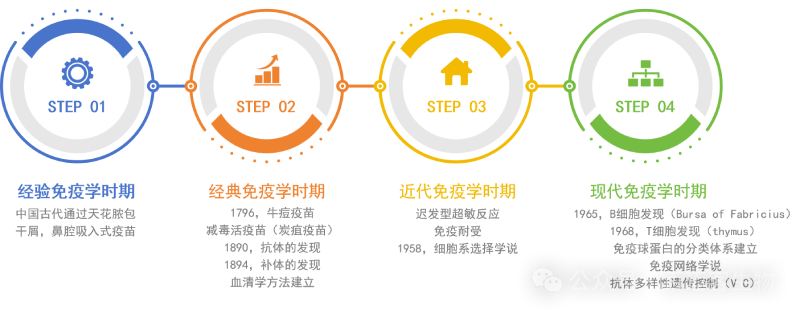
這是最早關(guān)于人類免疫學(xué)應(yīng)用的記錄。公元前?400?年,中國可能已有人痘苗接種預(yù)防天花的方法,到?16?世紀(jì)明朝隆慶年間,人痘接種法得到重大改進并廣泛應(yīng)用,17?世紀(jì)傳入俄國、朝鮮、日本、土耳其和英國等國家,至18世紀(jì)末結(jié)束經(jīng)驗免疫學(xué)時期,隨后免疫學(xué)又經(jīng)歷了經(jīng)典免疫學(xué)時期,近代免疫學(xué)時期,直到1965年/1968年?B細(xì)胞/T細(xì)胞的發(fā)現(xiàn)開啟了現(xiàn)代免疫學(xué)時期。
隨著免疫學(xué)的逐步發(fā)展,免疫組學(xué)技術(shù)也隨之進入了迅速發(fā)展的時期,先后發(fā)展出了標(biāo)記免疫組技術(shù)、免疫組化技術(shù)、Bulk?T/BCR測序技術(shù)以及近幾年興起的單細(xì)胞T/BCR測序技術(shù),并帶動了相關(guān)科學(xué)的進步。
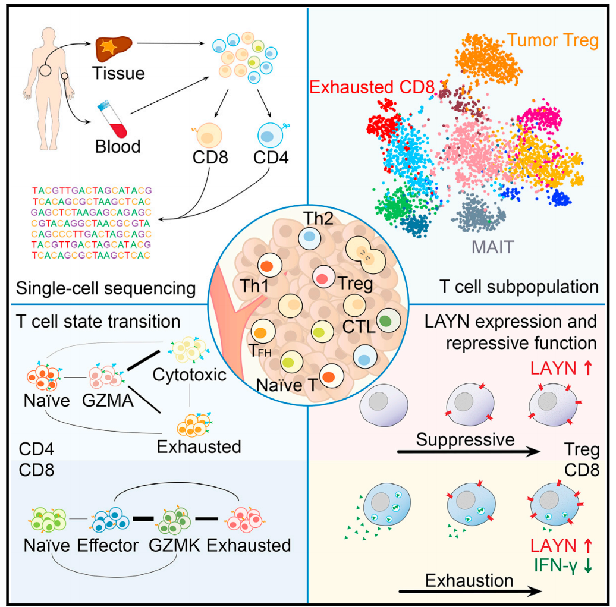
2017年,張澤民院士研究組,通過大規(guī)模單細(xì)胞測序技術(shù)對肝癌相關(guān)T淋巴細(xì)胞進行分析,首次在單細(xì)胞水平上描繪肝癌微環(huán)境中免疫細(xì)胞圖譜,發(fā)現(xiàn)了肝癌免疫療法的潛在靶點,也為我們從多角度系統(tǒng)性理解肝癌T淋巴細(xì)胞特征奠定了基礎(chǔ)。(2017,Cell,Landscape of Infiltrating T Cells in Liver Cancer Revealed by Single-Cell Sequencing)
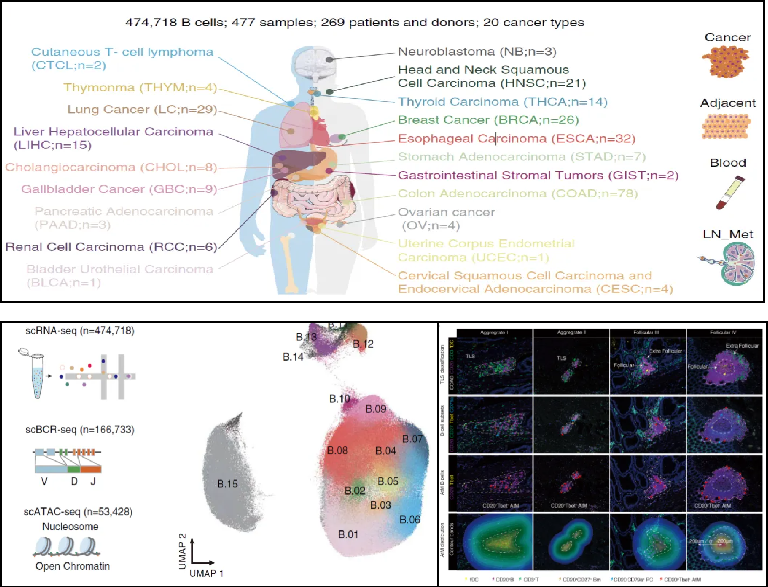
2024年,復(fù)旦大學(xué)附屬中山醫(yī)院樊嘉院士和高強教授、中國科學(xué)院上海免疫與感染研究所張曉明研究員、浙江大學(xué)基礎(chǔ)醫(yī)學(xué)院郭國驥教授合作在Science上發(fā)表了題為A blueprint for tumor-infiltrating B cells across human cancers的研究論文,結(jié)合單細(xì)胞轉(zhuǎn)錄組、 B細(xì)胞受體免疫組庫和表觀基因組的多組學(xué)數(shù)據(jù),系統(tǒng)性地刻畫了腫瘤浸潤性B細(xì)胞的異質(zhì)性、動態(tài)分化和表觀調(diào)控機制。創(chuàng)新地揭示了腫瘤微環(huán)境中廣泛存在的EF應(yīng)答的癌種偏好性、空間定位特征、臨床意義及潛在的誘導(dǎo)調(diào)控機制。
但是,與逐漸成熟的單細(xì)胞T/BCR測序不同,空間T/BCR測序測序技術(shù)還一直未能有較好的測序技術(shù)產(chǎn)品去做支撐,成果尚少,現(xiàn)有的文章技術(shù)還局限于使用早期的低分辨率轉(zhuǎn)錄芯片為依托。成熟穩(wěn)定且高分辨率的空間T/BCR測序技術(shù)產(chǎn)品的缺乏限制了空間免疫組的研究。
百創(chuàng)空間全長免疫組庫測序技術(shù)?BMKMANU?S3000-VDJ
2024年12月,百邁客生物智能制造以廣受市場好評的亞細(xì)胞級S系列空間轉(zhuǎn)錄組芯片為依托,開發(fā)出了在原片上可同時捕獲3’端轉(zhuǎn)錄組信息以及全長B細(xì)胞受體(BCR)和T細(xì)胞受體(TCR)序列的空間T/BCR測序技術(shù)產(chǎn)品—BMKMANU?S3000-VDJ?,將填補這一技術(shù)產(chǎn)品的缺口。
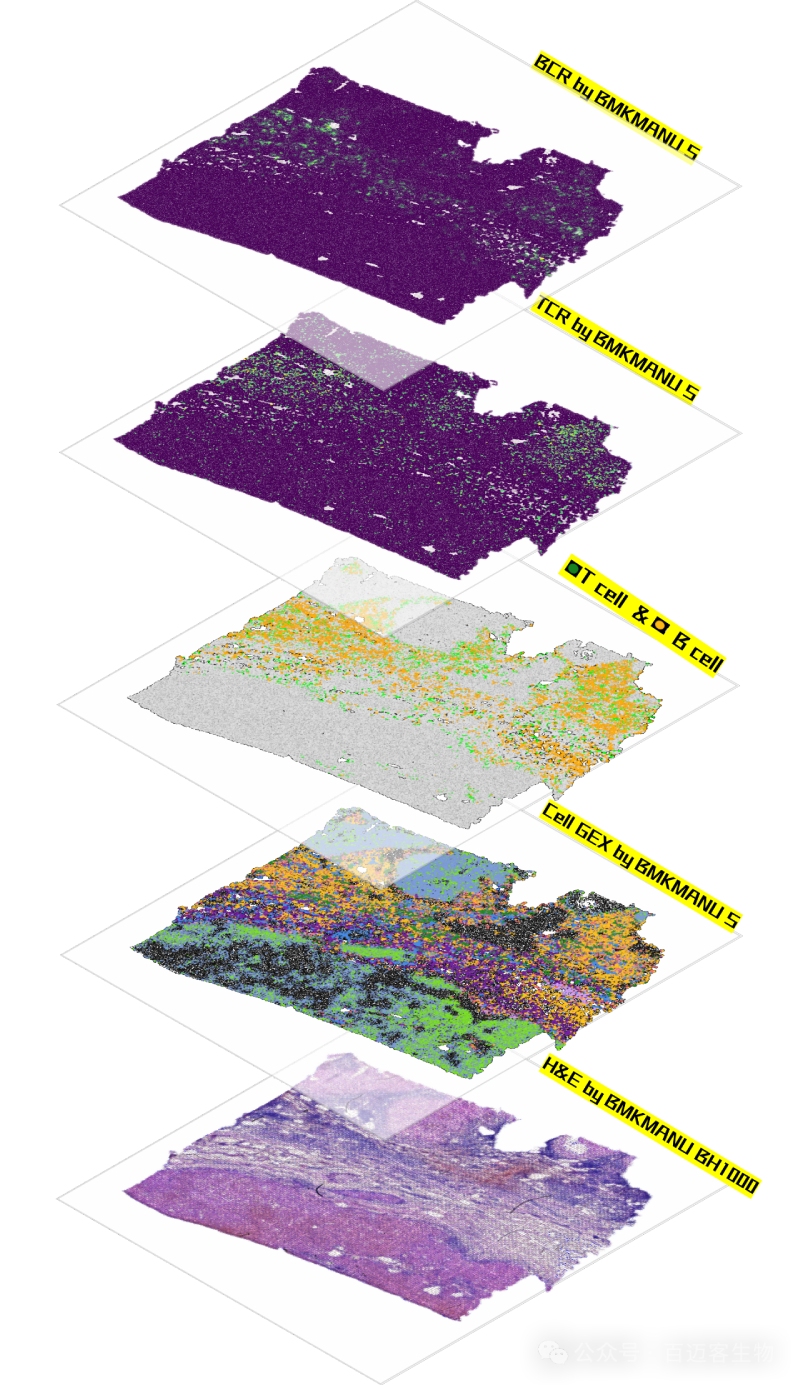
實測案例-某腫瘤-百創(chuàng)單細(xì)胞級空間免疫組結(jié)果
BMKMANU?S3000-VDJ?技術(shù),使用新鮮OCT包埋樣本,在分辨率為3.5μm的芯片上同一張組織切片(10μm)實現(xiàn)3’端mRNA,以及全長TCR/BCR的捕獲。利用百邁客生物智能制造獨有的圖像校準(zhǔn)及細(xì)胞分割技術(shù),得到單細(xì)胞級分辨率的空間轉(zhuǎn)錄組結(jié)果及單細(xì)胞級分辨率的空間T/BCR數(shù)據(jù)。
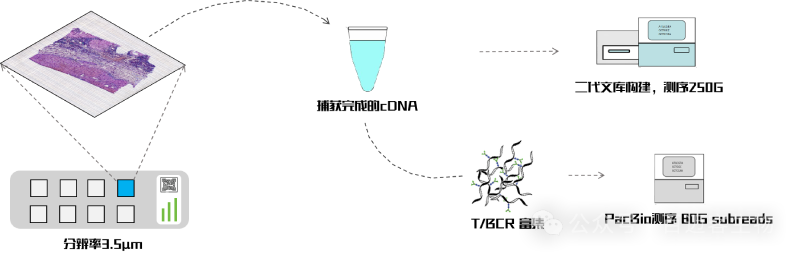
BMKMANU?S3000-VDJ??技術(shù)路線
精準(zhǔn)空間細(xì)胞注釋
得到的單細(xì)胞級空間轉(zhuǎn)錄組結(jié)果,可以進行常規(guī)的空間轉(zhuǎn)錄組分析,并進行精準(zhǔn)的細(xì)胞注釋。

精準(zhǔn)細(xì)胞注釋及T/B細(xì)胞的空間分布
T/B細(xì)胞的亞群定位
對于得到的T細(xì)胞及B細(xì)胞又可以繼續(xù)進行詳細(xì)的亞群分類,同時探討亞群之前的相互轉(zhuǎn)換關(guān)系以及與表型之前的關(guān)系。
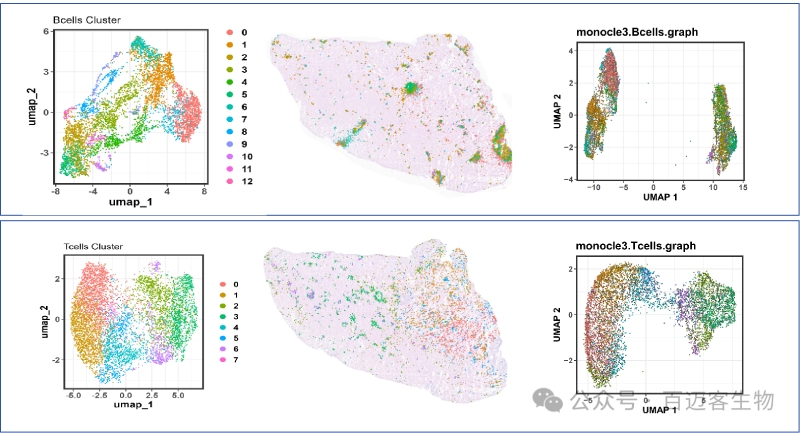
T/B Cells 亞群分類,亞群空間分布及亞群發(fā)育軌跡分析
TCR/BCR?空間分布以及豐度分析

TCR和BCR的豐度分析可以揭示免疫反應(yīng)的多樣性和動態(tài)變化。例如,TCR的多樣性在腫瘤進展過程中會發(fā)生變化,早期肝癌患者的TCR和BCR均勻性更高,而晚期患者則表現(xiàn)出較低的均勻性。此外,TCR和BCR的豐度變化也可以反映免疫細(xì)胞的激活狀態(tài)和克隆擴增情況。
克隆型空間分布以及豐度分析

腫瘤樣本我們可以類比為一處具有復(fù)雜地貌的特殊地理環(huán)境,不同的環(huán)境造就了各種細(xì)胞的不同狀態(tài),而T/B細(xì)胞在不同的“選擇壓力”下邊進行了不同的“適應(yīng)性進化”。對不同生態(tài)位的T/B細(xì)胞尤其是TCR/BCR進行測定,有助于我們?nèi)ソ沂久庖呒?xì)胞的克隆起源和進化過程。例如,通過分析TCR的VDJ重排,可以追蹤T細(xì)胞克隆的起源和擴增過程。這種分析有助于理解免疫細(xì)胞在不同生理和病理狀態(tài)下的動態(tài)變化。

百創(chuàng)單細(xì)胞級空間全長免疫組產(chǎn)品?BMKMANU?S3000-VDJ,將幫助科研者解決整個轉(zhuǎn)錄組和組織形態(tài)學(xué)問題,實現(xiàn)人類腫瘤或其它病變組織中B細(xì)胞和T細(xì)胞克隆的高保真定位和空間譜系追蹤,將在一定程度上促進對各種臨床相關(guān)現(xiàn)象(如感染、疫苗接種和癌癥)中淋巴細(xì)胞空間動力學(xué)的理解。
]]>百邁客生物為該研究提供了百創(chuàng)S1000空間轉(zhuǎn)錄組測序服務(wù)。

單細(xì)胞組學(xué)和時空組學(xué)是近年來生命科學(xué)領(lǐng)域的重要技術(shù)突破,它們在細(xì)胞異質(zhì)性解析、動態(tài)過程研究和細(xì)胞間相互作用分析方面具有重要優(yōu)勢,為生命科學(xué)研究帶來了全新的視角和深度。水稻稻瘟病是世界上發(fā)生嚴(yán)重的真菌病害之一,水稻在面臨稻瘟菌侵染時會激活動態(tài)免疫,因此挖掘水稻特異免疫基因?qū)τ诳沟疚敛【秩局陵P(guān)重要。
該研究對稻瘟菌接種后三個時間點的樣品進行了單細(xì)胞核轉(zhuǎn)錄組測序(圖1A),同時對接種后的水稻葉片也進了時空組測序(圖1B)。
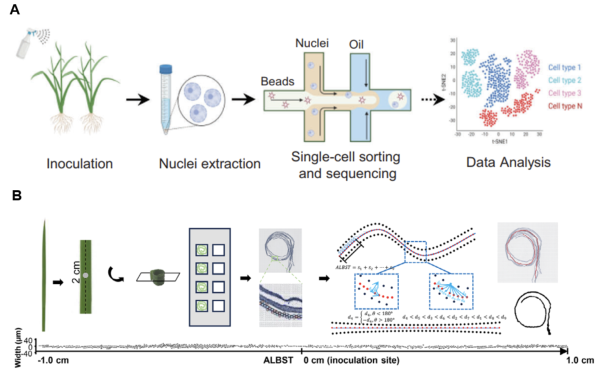
圖1?水稻樣品單細(xì)胞及時空組測序流程圖
對測序獲得的單細(xì)胞數(shù)據(jù),結(jié)合細(xì)胞特異的標(biāo)志基因,AddModuleScore?軟件和GO富集分析等方法,對水稻的細(xì)胞類型進行了分類注釋(圖2A)。基因表達分析發(fā)現(xiàn),維管細(xì)胞中的原形成層細(xì)胞在稻瘟菌侵染后高度誘導(dǎo)二萜類植保素合成代謝相關(guān)基因的表達,其中水稻關(guān)鍵植保素momilactone A合成被特異誘導(dǎo)(圖2B),促進了momilactone A在維管組織中積累,可能是抗稻瘟病的關(guān)鍵因素。相反,激素途徑和模式分子激發(fā)的免疫對抗病性貢獻較弱。
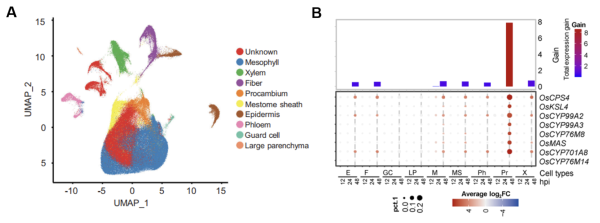
圖2?水稻單細(xì)胞測序結(jié)果及差異基因表達分析
共聚焦顯微鏡觀察結(jié)果顯示,稻瘟菌孢子萌發(fā)后優(yōu)先靶向葉脈入侵,但是水稻的維管組織具有較強的免疫,稻瘟菌菌絲無法進一步侵入(圖3A)。時空組測序結(jié)果顯示,富含亮氨酸重復(fù)序列的(NLR)免疫基因的表達在接種24小時后,葉尖部較葉基部具有更強的積累,表明免疫基因的表達具有極性的特征。綜上所述,該項研究揭示了水稻在稻瘟病菌侵染時免疫基因存在細(xì)胞特異性和多維(局部和縱向)的表達模式,為挖掘新的免疫基因提供了重要的數(shù)據(jù)基礎(chǔ)(圖3B)。
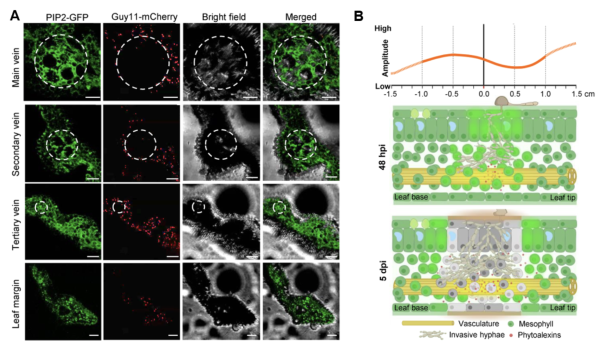
圖3 ?共聚焦顯微鏡觀察及時空組測序結(jié)果顯示水稻組織存在多維度的免疫
內(nèi)容來源于中國農(nóng)業(yè)大學(xué),侵刪
]]>
 該研究將空間轉(zhuǎn)錄組技術(shù)應(yīng)用于小麥籽粒發(fā)育研究,根據(jù)基因表達和位置信息詳細(xì)劃分細(xì)胞功能亞群,揭示了小麥籽粒發(fā)育過程的空間轉(zhuǎn)錄特征,繪制了小麥籽粒三維基因表達圖譜。
該研究將空間轉(zhuǎn)錄組技術(shù)應(yīng)用于小麥籽粒發(fā)育研究,根據(jù)基因表達和位置信息詳細(xì)劃分細(xì)胞功能亞群,揭示了小麥籽粒發(fā)育過程的空間轉(zhuǎn)錄特征,繪制了小麥籽粒三維基因表達圖譜。
百邁客生物為該研究提供了空間轉(zhuǎn)錄組測序服務(wù)。
小麥(Triticum aestivum L.)是全球三大主糧作物之一,種植面積約2.3億公頃,其產(chǎn)量提升對保障全球糧食安全與營養(yǎng)供給至關(guān)重要。小麥籽粒主要由三部分組成:二倍體胚、三倍體胚乳及果皮(種皮)。這些組織在發(fā)育過程中表現(xiàn)出顯著的基因表達時空特異性,形成功能分化的區(qū)室化結(jié)構(gòu)。盡管這些組織在細(xì)胞類型和生理功能上存在明顯差異,但它們通過協(xié)同調(diào)控構(gòu)建了獨特的籽粒發(fā)育微環(huán)境,共同決定籽粒的最終表型。然而,目前對籽粒細(xì)胞類型的認(rèn)知仍主要依賴于形態(tài)學(xué)觀察和少量標(biāo)記基因的表達分析,常規(guī)轉(zhuǎn)錄組數(shù)據(jù)因缺乏空間分辨率,極大限制了對籽粒發(fā)育調(diào)控機制的深入解析及關(guān)鍵功能基因的挖掘。
為系統(tǒng)解析小麥籽粒發(fā)育的分子機制,該研究采用時空轉(zhuǎn)錄組學(xué)技術(shù),構(gòu)建了籽粒在發(fā)育早期4 DAP、8 DAP和12 DAP的高分辨率基因表達圖譜,實現(xiàn)了近8萬個基因的空間表達可視化,并鑒定了10個特征明確的細(xì)胞亞群及190個籽粒特異性標(biāo)記基因。研究發(fā)現(xiàn),不同組織特異性基因(尤其是轉(zhuǎn)錄因子)呈現(xiàn)動態(tài)表達模式。通過共表達網(wǎng)絡(luò)和順式調(diào)控元件分析,鑒定到一個關(guān)鍵調(diào)控因子——轉(zhuǎn)錄因子TaABI3-B1,其在胚胎及胚乳周圍組織中特異性表達,并負(fù)向調(diào)控胚胎和籽粒大小。進一步利用等位基因變異體和轉(zhuǎn)基因材料驗證了TaABI3-B1在胚胎和胚乳發(fā)育中的關(guān)鍵作用。

該研究不僅為小麥籽粒發(fā)育提供了全面的時空轉(zhuǎn)錄組資源,還揭示了調(diào)控籽粒大小的新機制,為小麥分子設(shè)計育種和產(chǎn)量提升提供了重要理論依據(jù)。
內(nèi)容來源于北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院,侵刪
]]>

通過對多組學(xué)數(shù)據(jù)的深度整合分析,該研究全面探究了重要木本經(jīng)濟植物核桃(Juglans regia)及其近緣種在基因組層面的遺傳變異特征。同時,深入剖析了核桃果實發(fā)育過程中的轉(zhuǎn)錄代謝調(diào)控機制,揭示了表觀遺傳修飾以及空間轉(zhuǎn)錄調(diào)控在其中的關(guān)鍵作用,并對相關(guān)候選基因的功能進行了系統(tǒng)分析。該研究為深入理解核桃果殼基因變異及其網(wǎng)絡(luò)遺傳調(diào)控機制提供了全新的視角和理論依據(jù),為推動核桃分子育種技術(shù)的創(chuàng)新發(fā)展奠定了堅實的基礎(chǔ)。
百邁客生物為該研究提供了植物空間轉(zhuǎn)錄組測序服務(wù)。
研究背景
核桃(Juglans regia)是一種具有重要經(jīng)濟價值的堅果油料樹種;其果實具有堅硬的內(nèi)果皮/果殼以保護種子,這在其進化過程中發(fā)揮了關(guān)鍵作用,而果殼厚度是核桃育種的一個重要性狀。然而,與核桃果殼發(fā)育相關(guān)的基因組景觀和基因調(diào)控網(wǎng)絡(luò)尚未得到系統(tǒng)的闡明。
研究內(nèi)容
該研究組裝注釋了核桃品種“香玲”的高質(zhì)量基因組,并構(gòu)建了涵蓋八個胡桃屬物種的圖形結(jié)構(gòu)泛基因組,以揭示基因組水平上的遺傳變異。重新測序了285份樣本,揭示了其群體基因組景觀變異圖譜。通過全基因組關(guān)聯(lián)研究(GWAS),研究發(fā)現(xiàn)了19個與核桃馴化和改良過程中受到選擇的268多個位點相關(guān)的基因座。

圖1-核桃比較基因組學(xué)與遺傳變異分析
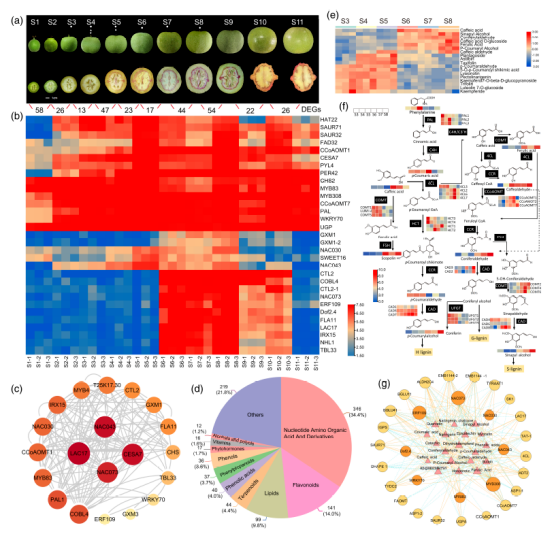
圖2-核桃果殼/內(nèi)果皮十一個發(fā)育階段基因轉(zhuǎn)錄調(diào)控網(wǎng)絡(luò)及動態(tài)形態(tài)變化
通過對十一個發(fā)育階段的轉(zhuǎn)錄組學(xué)、代謝組學(xué)、DNA甲基化和空間轉(zhuǎn)錄組學(xué)等多組學(xué)分析,揭示了多個與次生細(xì)胞生物合成和木質(zhì)素積累相關(guān)的候選基因,例如UGP、MYB308、MYB83、NAC043、NAC073、CCoAOMT1、CCoAOMT7、CHS2、CESA7、LAC7、COBL4和IRX12。在擬南芥(Arabidopsis thaliana)中過表達JrUGP和JrMYB308驗證了它們在木質(zhì)素生物合成和細(xì)胞壁加厚中的作用。

研究結(jié)論
植物的果實特征,包括核桃(Juglans)的堅硬內(nèi)果皮,在種子傳播、農(nóng)業(yè)生產(chǎn)效率和遺傳多樣性方面發(fā)揮著至關(guān)重要的作用,其中殼厚對核桃的生產(chǎn)和育種有著顯著影響。綜合多組學(xué)分析探究了核桃殼厚度這一關(guān)鍵性狀以及與木質(zhì)素積累和細(xì)胞壁增厚相關(guān)的內(nèi)果皮發(fā)育,為木本作物的遺傳學(xué)和調(diào)控機制提供了新的見解,助力基于基因組信息的改良育種策略。全面多組學(xué)研究結(jié)果為核桃的遺傳變異和內(nèi)果皮發(fā)育及果殼厚度的網(wǎng)絡(luò)調(diào)控提供了新的見解,并為核桃品種改良的基因組信息育種策略提供了可能。
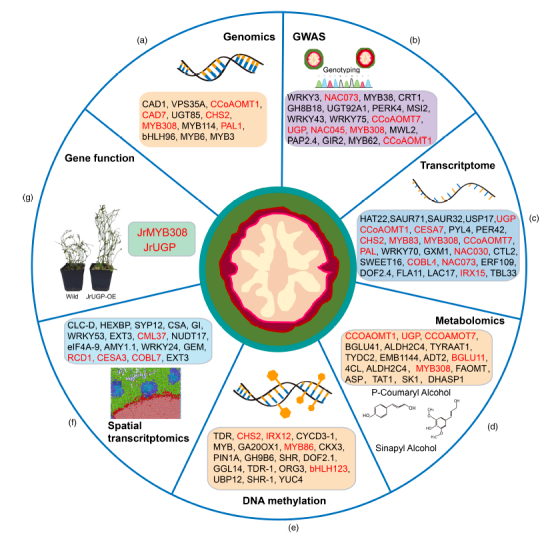
圖4-該研究主要發(fā)現(xiàn)概括模型
內(nèi)容來源于西北大學(xué)生命科學(xué)學(xué)院,侵刪
]]>

文章標(biāo)題:Spatial and single-cell transcriptomics reveal cellular heterogeneity and a novel cancer-promoting Treg cell subset in human clear-cell renal cell carcinoma
期刊名稱:Journal for ImmunoTherapy of Cancer
影響因子:10.3
合作單位:空軍軍醫(yī)大學(xué)(第四軍醫(yī)大學(xué))
研究方法:單細(xì)胞轉(zhuǎn)錄組測序,空間轉(zhuǎn)錄組測序,流式分選,RT-qPCR,Western blot,免疫組化,多重免疫組化,細(xì)胞實驗。
百邁客生物為該研究提供了百創(chuàng)DG1000單細(xì)胞轉(zhuǎn)錄組測序服務(wù)以及百創(chuàng)S1000空間轉(zhuǎn)錄組技術(shù)服務(wù),文中百創(chuàng)S1000空間數(shù)據(jù)分析得到的細(xì)胞空間分布圖由百創(chuàng)開發(fā)軟件BSTCellViewer展示。
研究背景
透明腎細(xì)胞癌ccRCC是腎細(xì)胞癌RCC的常見亞型,是腎癌相關(guān)死亡的主要原因,治療手段包括腎切除手術(shù)、靶向治療和免疫治療,但多數(shù)患者不能從免疫治療獲益,這與腫瘤微環(huán)境異質(zhì)性有關(guān),因此有必要深入理解ccRCC的腫瘤微環(huán)境,助力開發(fā)新型個性化診療方案。
材料及方法
研究材料:15名腎切除術(shù)的ccRCC患者組織樣本及其血液PBMC。對組織樣本區(qū)域進行定義,分為腫瘤核心(TC),腫瘤-健康交界(IF,包括腫瘤邊緣TR和臨近健康腎組織AN),遠(yuǎn)端健康腎組織DN。
組別設(shè)計:實驗組4名ccRCC患者樣本進行單細(xì)胞轉(zhuǎn)錄組測序&空間轉(zhuǎn)錄組測序,驗證組15名ccRCC患者樣本進行流式細(xì)胞術(shù)&多重免疫熒光染色,151名ccRCC患者組織樣本&40名其他腎癌亞型患者組織樣本&6名捐獻者健康腎組織進行多重?zé)晒饷庖呓M化染色。
研究方法:單細(xì)胞轉(zhuǎn)錄組測序(4名患者不同區(qū)域腫瘤組織,按照CD45+細(xì)胞:CD45-細(xì)胞=9:1增加免疫細(xì)胞數(shù)據(jù)占比,n=12),空間轉(zhuǎn)錄組測序(n=4);流式分選,RT-qPCR,Western blot,免疫組化,多重免疫組化,細(xì)胞實驗。
研究結(jié)果
1.ccRCC組織細(xì)胞類型全面分析
單細(xì)胞轉(zhuǎn)錄組測序(scRNA-seq)數(shù)據(jù)顯示不同患者腫瘤組織的免疫微環(huán)境細(xì)胞組成相似,T細(xì)胞占比最高,特別是CD4+ T細(xì)胞。腫瘤區(qū)域(TC/TR)富集CD3+細(xì)胞以及CD8+ T細(xì)胞,而CD4+ T細(xì)胞基本分布在所有區(qū)域。此外,基質(zhì)細(xì)胞和免疫細(xì)胞豐度和分布存在差異,TR區(qū)域成纖維細(xì)胞比例顯著低于TC區(qū)域和AN區(qū)域,這一發(fā)現(xiàn)也在空間轉(zhuǎn)錄數(shù)據(jù)中得到印證。
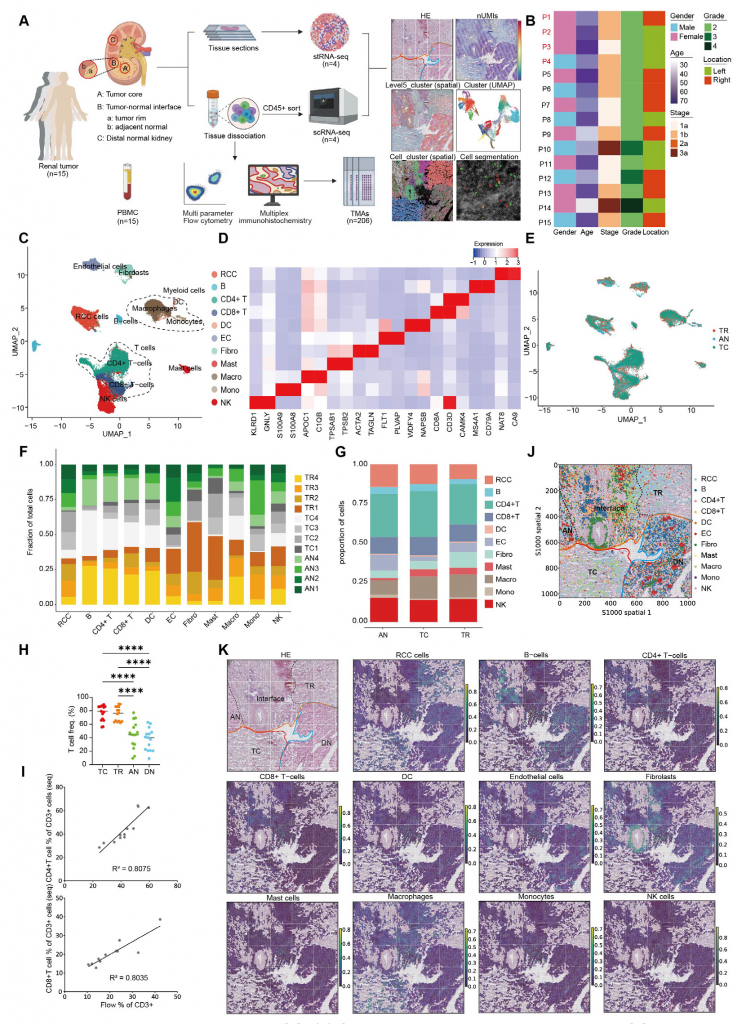
圖1-ccRCC組織中細(xì)胞群的全面分析
2.ccRCC細(xì)胞元程序的豐度以及異質(zhì)性
通過非負(fù)矩陣因子分解分析每個樣本scRNA-seq數(shù)據(jù),得到可以代表ccRCC腫瘤細(xì)胞簇元程序的6個基因集,不同基因集代表著不同的程序,如基因集1代表著EMT(上皮-間充質(zhì)轉(zhuǎn)變)相關(guān)基因表達譜,基因集5也與EMT相關(guān)但與基因集1不同的是其包含VEGFA等。
進一步分析,發(fā)現(xiàn)ccRCC亞群0~4簇與不同基因集表達譜匹配,5和6簇不與任何基因集匹配但具有細(xì)胞周期和應(yīng)激響應(yīng)相關(guān)基因表達。RCC0簇和RCC2簇代表EMThigh腫瘤細(xì)胞并在IF區(qū)域富集,RCC1簇代表PT(近曲小管)細(xì)胞,擬時序分析結(jié)果顯示存在RCC0—>RCC2分化關(guān)系,但RCC2幾乎只在P2患者樣本中發(fā)現(xiàn),表明RCC2可能與個別患者特征相關(guān)。RCC1、RCC3和RCC4細(xì)胞主要在組織而不是腫瘤區(qū)域富集,RCC4主要分布在AN和DN區(qū)域,RCC4在AN區(qū)域分布數(shù)量高于DN區(qū)域。RCC4高表達細(xì)胞因子受體基因,表明TME中分泌的促腫瘤細(xì)胞因子可能傾向于作用到RCC4,增加其轉(zhuǎn)變成癌細(xì)胞的概率,加速腫瘤逃逸,促進腫瘤惡性化,與以往瘤旁附近不是完全的健康組織認(rèn)識相符合。
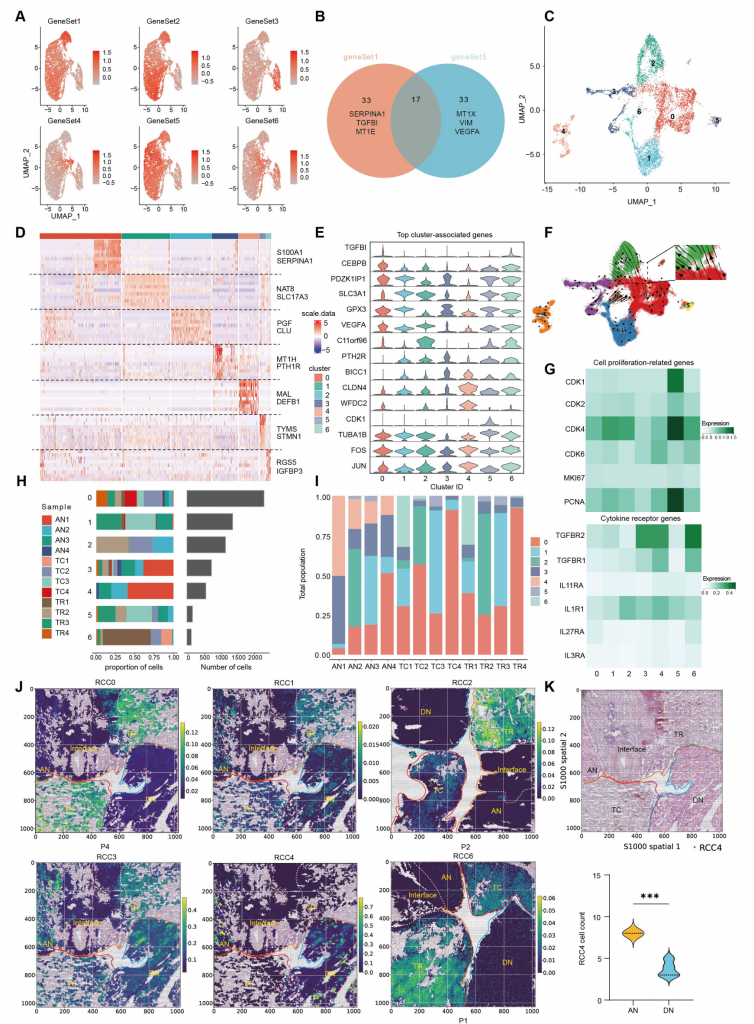
圖2-透明細(xì)胞RCC細(xì)胞元程序的豐度及異質(zhì)性
3.ccRCC基質(zhì)細(xì)胞的區(qū)域化特征和異質(zhì)性
scRNA-seq與空間數(shù)據(jù)均顯示,ccRCC存在8種激活的成纖維細(xì)胞簇,腫瘤區(qū)域成纖維細(xì)胞數(shù)量高于健康組織,其中CD24+IL32+成纖維細(xì)胞顯著富集在腫瘤區(qū)域,PTGDS+成纖維細(xì)胞幾乎全分布在AN,高表達ECM(胞外基質(zhì))形成和EMT的成纖維細(xì)胞簇在IF較為富集,IL1R1+?CAF(腫瘤相關(guān)成纖維細(xì)胞)表現(xiàn)出腫瘤促進表型;7種EC(內(nèi)皮細(xì)胞)簇中4種(0,2,3和6)主要富集在腫瘤組織,3種(1,4和5)在健康組織富集,膠原蛋白EC(0簇)豐度最高,具有促腫瘤特征。總之,不同的ECM蛋白質(zhì)產(chǎn)生基質(zhì)細(xì)胞傾向于富集在IF并共定位,可能行駛多種功能,包括細(xì)胞-細(xì)胞交互和胞外組分重構(gòu)。

圖3-透明細(xì)胞腎細(xì)胞癌中基質(zhì)細(xì)胞的區(qū)域特征和異質(zhì)性
4.ccRCC中MRC1+FOLR2+?TAM傾向于表現(xiàn)出更強的促癌表型
scRNA-seq數(shù)據(jù)與空間數(shù)據(jù)均顯示,4種TAM簇具有不同分布趨勢,TAM1主要分布在TC區(qū)域;TAM2高表達FOLR2、MRC1和CD163,可能是M2樣巨噬細(xì)胞且具有增強的免疫抑制特征,TAM4是組織駐留巨噬細(xì)胞,兩者主要分布在TR區(qū)域;TAM3表達炎癥響應(yīng)相關(guān)基因,在TR和AN區(qū)域富集。RNA速率分析顯示存在TAM4—>TAM2—>TAM1和TAM4—>TAM3兩種軌跡,以及從IF區(qū)域遷移到TC區(qū)域的趨勢。進一步的功能分析顯示,IF區(qū)域的TAM2可能通過促進EMT,TC區(qū)域的TAM2可能通過促進腫瘤血管生成,起到促癌效應(yīng)。總之,TAM2不同的腫瘤促進效應(yīng)可能與免疫抑制TME有關(guān)。

圖4-透明細(xì)胞腎細(xì)胞癌中MRC1+FOLR2+?TAM傾向于表現(xiàn)更強的促癌表型
5.單細(xì)胞分析揭示ccRCC中存在表達IL1B的Treg
進一步分析CD4+ T細(xì)胞簇,發(fā)現(xiàn)存在CD4+ 初始T細(xì)胞、Treg和濾泡樣輔助T細(xì)胞,不同簇的特征基因和空間分布模式不同,IF區(qū)域富集CD4+ 初始T細(xì)胞和PDE3B+ CD4+ T細(xì)胞,腫瘤區(qū)域富集Treg。其中,TC/TR區(qū)域CTLA-4+?Treg的占比以及Treg的FoxP3蛋白表達水平高于AN/DN區(qū)域,而CTLA-4+FoxP3-?Treg在所有組織中的分布較為均勻,表明腫瘤區(qū)域Treg細(xì)胞具有更強的免疫抑制功能。
進一步分析Treg細(xì)胞簇,發(fā)現(xiàn)存在6種亞型,其中終末效應(yīng)Treg細(xì)胞(5)主要分布在IF區(qū)域,F(xiàn)OXP3表達水平較低且ICOS表達水平降低,表明細(xì)胞可能處于不穩(wěn)定狀態(tài)。
此外,終末效應(yīng)Treg細(xì)胞表達炎癥性細(xì)胞因子如IL1B,IL18和IL7,驗證隊列的多重免疫熒光染色結(jié)果顯示IL-1β+終末效應(yīng)Treg是對ccRCC特異性腫瘤微環(huán)境信號的特異性響應(yīng)。發(fā)現(xiàn)腫瘤組織中存在表達炎癥性細(xì)胞因子IL1B的終末效應(yīng)Treg細(xì)胞。

圖5-單細(xì)胞分析揭示透明細(xì)胞腎細(xì)胞癌存在表達IL1B的Treg細(xì)胞群
6.IL-18通過ERK/NF-κB通路促進強免疫抑制功能的終末效應(yīng)Treg細(xì)胞產(chǎn)生
空間轉(zhuǎn)錄組數(shù)據(jù)結(jié)果顯示不同Treg細(xì)胞100μm范圍內(nèi)激活的CD4+/CD8+ T細(xì)胞數(shù)量較少,終末效應(yīng)Treg細(xì)胞100μm范圍內(nèi)CD4+/CD8+ T細(xì)胞數(shù)量更少,表明終末效應(yīng)Treg細(xì)胞周圍應(yīng)答Treg細(xì)胞(Tresp)增殖受到更強的抑制。細(xì)胞共培養(yǎng)實驗表明終末效應(yīng)Treg細(xì)胞具有強免疫抑制功能。RNA速率分析結(jié)果表明效應(yīng)終末Treg可能由初始Treg細(xì)胞分化而來,體外實驗表明IL-18可刺激CD4+CD25+ Treg細(xì)胞上調(diào)IL-1β表達,表明終末效應(yīng)Treg細(xì)胞產(chǎn)生和作用可能與IL-18有關(guān)。ERK激活劑/NF-κB激活劑促進IL-18+ Treg細(xì)胞的產(chǎn)生,IL-18處理后Treg細(xì)胞ERK/NF-κB磷酸化水平增加,ERK抑制劑可抑制Treg細(xì)胞表達NF-κB但NF-κB抑制劑無法抑制ERK表達,綜上,這些結(jié)果表明IL-18可能通過ERK/NF-κB通路介導(dǎo)終末效應(yīng)Treg細(xì)胞的產(chǎn)生。

圖6-IL-18通過ERK/NF-κB通路促進具有強免疫抑制功能的終末效應(yīng)Treg細(xì)胞產(chǎn)生
7.通過與MRC1+FLOR2+?TAM互作,終末效應(yīng)Treg細(xì)胞與存活率降低、免疫逃逸增加及腫瘤生長相關(guān)
確定終末效應(yīng)Treg細(xì)胞標(biāo)志基因(FOXP3、IL1B和FCER1G),TGCA泛癌數(shù)據(jù)庫數(shù)據(jù)分析結(jié)果顯示,終末效應(yīng)T細(xì)胞浸潤高的隊列與更低的整體存活率相關(guān)。細(xì)胞通訊分析結(jié)果顯示,巨噬細(xì)胞與終末效應(yīng)Treg細(xì)胞有著最多的互作配體-受體對,許多是免疫抑制互作,驗證實驗支持終末效應(yīng)Treg細(xì)胞與TAM2簇之間存在相互作用;IL1R1+?CAF與終末效應(yīng)Treg細(xì)胞也存在潛在聯(lián)系。僅在TC區(qū)域(與TR區(qū)域相比)發(fā)現(xiàn)NAMPT-INSR和TGFB1-(TGFBR1+TGFBR2)配受體互作,體外實驗發(fā)現(xiàn)終末效應(yīng)Treg細(xì)胞LRRC32(該基因編碼的GARP是TGF-β1成熟和激活所必須的)表達水平高于傳統(tǒng)Treg細(xì)胞。
此外,巨噬細(xì)胞特別是TAM2高表達IL18。這些結(jié)果可能表明ccRCC腫瘤微環(huán)境中,IF區(qū)域終末效應(yīng)Treg細(xì)胞與臨近TAM2互作,通過分泌的TGF-β1、M-CSF1以及IL-10,將IF駐留的TAM轉(zhuǎn)變成促癌表型,引起腫瘤細(xì)胞EMT;TAM2分泌的趨化因子誘導(dǎo)Treg細(xì)胞遷移到腫瘤區(qū)域并過表達IL-18,將Treg細(xì)胞轉(zhuǎn)變成IL-1β+ 終末效應(yīng)T細(xì)胞,抑制T細(xì)胞免疫并促進腫瘤生長。
總之,這些結(jié)果表明ccRCC的IF區(qū)域,終末效應(yīng)Treg細(xì)胞可能與其他促癌細(xì)胞互作,最終促進腫瘤細(xì)胞惡性轉(zhuǎn)化,支持了終末效應(yīng)Treg細(xì)胞具有免疫抑制和促癌作用的假設(shè)。
研究結(jié)果
該研究系統(tǒng)描述了ccRCC中不同類型細(xì)胞的基因表達譜和空間分布規(guī)律,發(fā)現(xiàn)一種具有癌癥促進作用的新Treg細(xì)胞亞型,在腫瘤-健康組織交界區(qū)域與MRC1+FOLR2+?TAM互作,構(gòu)建免疫抑制TME,促進腫瘤細(xì)胞惡性轉(zhuǎn)化最終引起患者存活率降低。同時,這些發(fā)現(xiàn)也為開發(fā)新的藥物和預(yù)后標(biāo)志物提供了靶點。
參考文獻:
Song X, et al. Spatial and single-cell transcriptomics reveal cellular heterogeneity and a novel cancer-promoting Treg cell subset in human clear-cell renal cell carcinoma. J Immunother Cancer. 2025 13(1):e010183.
]]>
2025年3月,華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院器官移植研究所蘭培祥教授、陳知水教授和肝臟外科程琪教授研究團隊在Journal of Hepatology發(fā)表題為”Sour Neuronal Signalling Attenuates Macrophage Mediated Liver Injury”的研究論文。研究使用單細(xì)胞轉(zhuǎn)錄組測序技術(shù)等多種技術(shù),深入闡述人鼠中減輕肝缺血再灌注損傷的腦-肝軸調(diào)控通路,發(fā)現(xiàn)酸味刺激竟然可以通過神經(jīng)信號緩解肝臟損傷!這一發(fā)現(xiàn)不僅為肝臟疾病的治療提供了新思路,還從現(xiàn)代科學(xué)的角度驗證了中醫(yī)“酸入肝”的理論。
文章標(biāo)題:Sour Neuronal Signalling Attenuates Macrophage Mediated Liver Injury
期刊名稱:Journal of Hepatology
影響因子:26.7
合作單位:華中科技大學(xué)同濟醫(yī)學(xué)院附屬同濟醫(yī)院
百邁客生物為該研究提供了10X單細(xì)胞轉(zhuǎn)錄組測序技術(shù)服務(wù)。
研究背景
在中醫(yī)理論中,酸味與肝臟有著密切的關(guān)系。中醫(yī)經(jīng)典《黃帝內(nèi)經(jīng)》中提到“酸入肝”,認(rèn)為酸味食物能夠滋養(yǎng)肝臟,調(diào)和氣血,促進肝臟的生理功能。像檸檬、山楂、醋等酸味食物,常被用來調(diào)理肝氣郁結(jié)、疏肝解郁。這次的研究,不僅讓中醫(yī)的古老智慧得到了科學(xué)驗證,還為酸味在肝臟疾病治療中的應(yīng)用提供了新的依據(jù)。
肝損傷是多種肝病常見的病理生理基礎(chǔ),與炎癥有關(guān)。肝缺血再灌注損傷是一種局部無菌炎癥響應(yīng),主要由先天免疫細(xì)胞驅(qū)動,是肝切除術(shù)中早期器官功能障礙和衰竭的重要原因。傳統(tǒng)認(rèn)為肝缺血再灌注引起的炎癥是由肝和死亡細(xì)胞釋放的DAMP(損傷相關(guān)分子蛋白)被肝駐留庫普夫細(xì)胞、單核細(xì)胞、單核-巨噬細(xì)胞等識別,釋放趨化因子和促炎癥因子,招募循環(huán)白細(xì)胞促使炎癥發(fā)生。此外,近年來,神經(jīng)免疫調(diào)控成為研究熱點,科學(xué)家們發(fā)現(xiàn),神經(jīng)系統(tǒng)可以通過釋放神經(jīng)遞質(zhì)、神經(jīng)肽等分子來調(diào)節(jié)免疫反應(yīng)。然而,如何通過神經(jīng)信號來治療肝臟炎癥,仍然是一個未解之謎。
材料及方法
研究方法:單細(xì)胞核轉(zhuǎn)錄組測序(n=3,肝及腹腔神經(jīng)節(jié)),組織學(xué)染色,熒光染色,病毒示蹤,免疫印記,免疫沉淀串聯(lián)質(zhì)譜,bulk RNA-seq,qRT-PCR,流式細(xì)胞術(shù)等。
研究材料:C57BL/6J小鼠缺血再灌注損傷模型,Fam19a2-/-Ccr2-/-小鼠,小鼠海馬神經(jīng)元細(xì)胞系HT122。
研究結(jié)果
1.酸刺激減輕肝缺血再灌注損傷
研究者首先構(gòu)建酸刺激下肝臟缺血再灌注損傷(IRI)小鼠模型,發(fā)現(xiàn)酸刺激可以減少肝組織損傷以及血清標(biāo)志物水平。但是,使用丁卡因局麻小鼠舌頭或者灌胃檸檬酸不能減少肝損傷和血清標(biāo)志物水平;NMDAR阻斷劑1(阻斷NMDAR介導(dǎo)的興奮性突觸傳遞)立體定位注射到腹后內(nèi)側(cè)核(VPN)后,酸刺激后IRI肝的血清標(biāo)志物水平和肝損傷程度無明顯變化,表明神經(jīng)系統(tǒng)在酸刺激減輕IRI過程中起重要作用。此外,小鼠VPN注射示蹤病毒的實驗結(jié)果顯示,肝臟以及CG(腹腔神經(jīng)節(jié))均檢測到熒光,行腹腔神經(jīng)節(jié)切除術(shù)能降低IRI肝的組織損傷和血清標(biāo)志物水平,表明酸刺激減輕肝損傷過程通過腦-CG-肝軸。

圖1-酸通過神經(jīng)減輕小鼠肝臟缺血再灌注損傷

圖2-腦和肝中分布的H129感染神經(jīng)
2.酸刺激通過減少TAFA2產(chǎn)生降低肝IRI
對肝IRI小鼠、酸刺激后肝IRI小鼠及sham(假手術(shù))小鼠的肝臟和CG進行單細(xì)胞核轉(zhuǎn)錄組測序,結(jié)果顯示IRI引起肝細(xì)胞和神經(jīng)元基因表達譜發(fā)生變化,IRI肝中免疫細(xì)胞數(shù)量增加,但酸刺激后IRI樣本中免疫細(xì)胞數(shù)量減少。神經(jīng)元較為特異性地表達TAFA2,IRI可引起肝神經(jīng)元TAFA2表達水平增加,但酸刺激使得TAFA2表達水平降到與sham對照組相近水平。實驗發(fā)現(xiàn)鉀離子可引起小鼠海馬神經(jīng)元細(xì)胞系HT22顯著高表達TAFA2,使用鉀離子通道抑制劑可減輕肝損傷,支持酸刺激通過神經(jīng)系統(tǒng)減輕肝損傷的假設(shè)。進一步的,使用慢病毒敲低神經(jīng)元TAFA2表達水平或使用TAFA2敲基因鼠,發(fā)現(xiàn)IRI引起的肝組織損傷、巨噬細(xì)胞浸潤以及血清標(biāo)志物水平降低,表明TAFA2在肝IRI中有著重要作用。
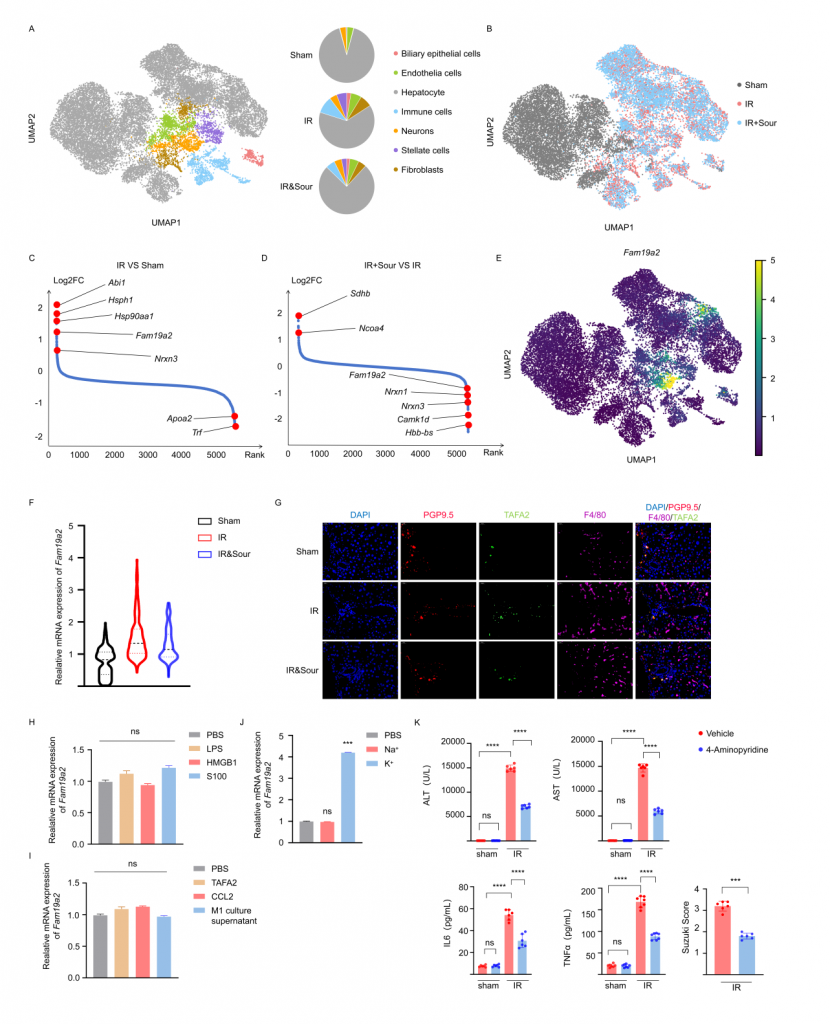
圖3-酸刺激減少小鼠IRI肝中神經(jīng)細(xì)胞Fam19a2表達水平

圖4-Fam19a2敲除或敲低,使得小鼠肝臟缺血再灌注損傷降低
?3.IRI肝中TAFA2與巨噬細(xì)胞相互作用
體外實驗發(fā)現(xiàn)TAFA2不引起肝細(xì)胞凋亡;snRNA-seq數(shù)據(jù)顯示IRI肝中巨噬細(xì)胞比例增加且炎癥相關(guān)基因表達水平增加,但在酸刺激組中降低;流式細(xì)胞術(shù)實驗結(jié)果表明,TAFA2與巨噬細(xì)胞結(jié)合而不與T/B細(xì)胞結(jié)合,不論是否酸刺激T/B細(xì)胞比例無顯著變化,IRI肝中CD11b+F4/80low?巨噬細(xì)胞增加而酸刺激可降低該巨噬細(xì)胞數(shù)量;TAFA2敲除或敲低抑制IRI肝中巨噬細(xì)胞的浸潤,這些結(jié)果表明TAFA2促進巨噬細(xì)胞活化。體外實驗結(jié)果表明,TAFA2可以激活BMDM(骨髓來源巨噬細(xì)胞),引起Il1α,Il1β,Il6和Tnfα的表達,這些結(jié)果表明TAFA2激活的巨噬細(xì)胞介導(dǎo)了肝IRI。
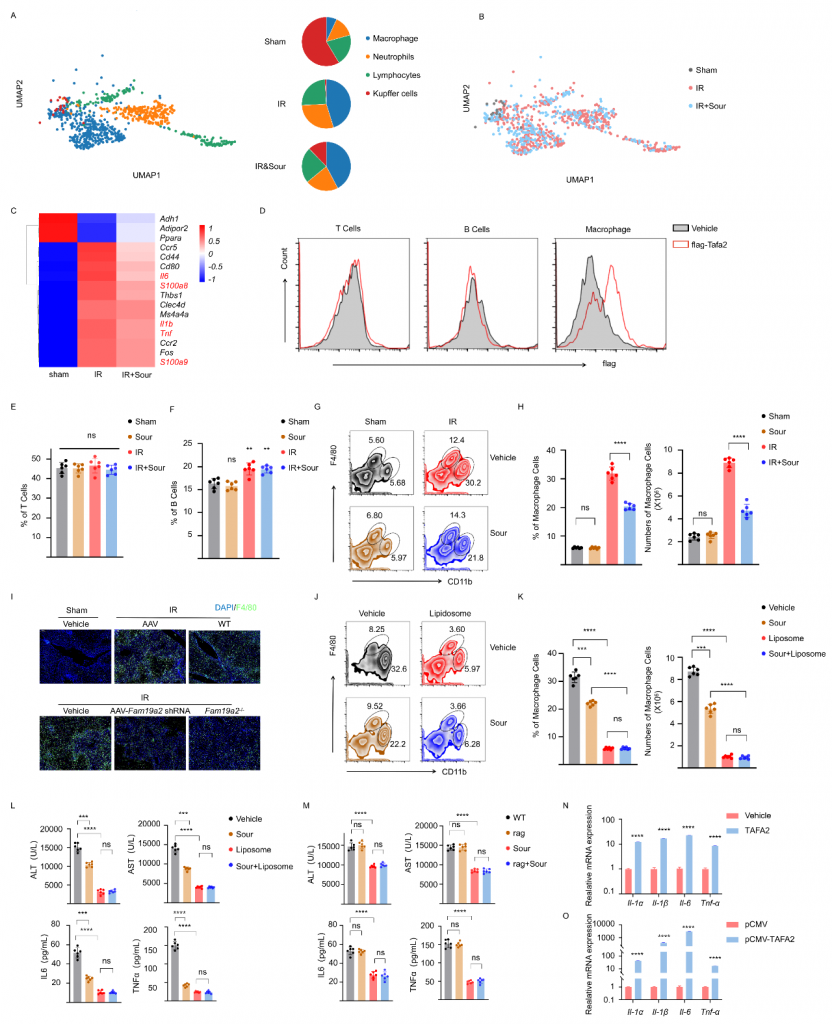
圖5-TAFA2使得IRI中巨噬細(xì)胞比例增加,刺激炎性細(xì)胞因子的產(chǎn)生
4.TAFA2通過CCR2與巨噬細(xì)胞互作
體外實驗表明巨噬細(xì)胞上的CCR2是TAFA2受體,CCR2敲除小鼠IRI肝的組織損傷以及巨噬細(xì)胞浸潤降低,血清標(biāo)志物水平降低。TAFA2或CCL2刺激后BMDM的bulk RNA-seq數(shù)據(jù)表現(xiàn)出幾乎一致的轉(zhuǎn)錄組表達譜,粘附、代謝、Ras信號通路相關(guān)差異基因表達上調(diào),代謝和核糖體通路相關(guān)基因表達下調(diào),TAFA2誘導(dǎo)BMDM更高的表達Il1α,Il1β,Il6,Tnfα以及干擾素相關(guān)基因,促進巨噬細(xì)胞介導(dǎo)的炎癥響應(yīng)。進一步實驗表明TAFA2促使巨噬細(xì)胞介導(dǎo)的炎癥響應(yīng)主要依賴CCR2,但巨噬細(xì)胞上也可能存在其他的TAFA2受體。
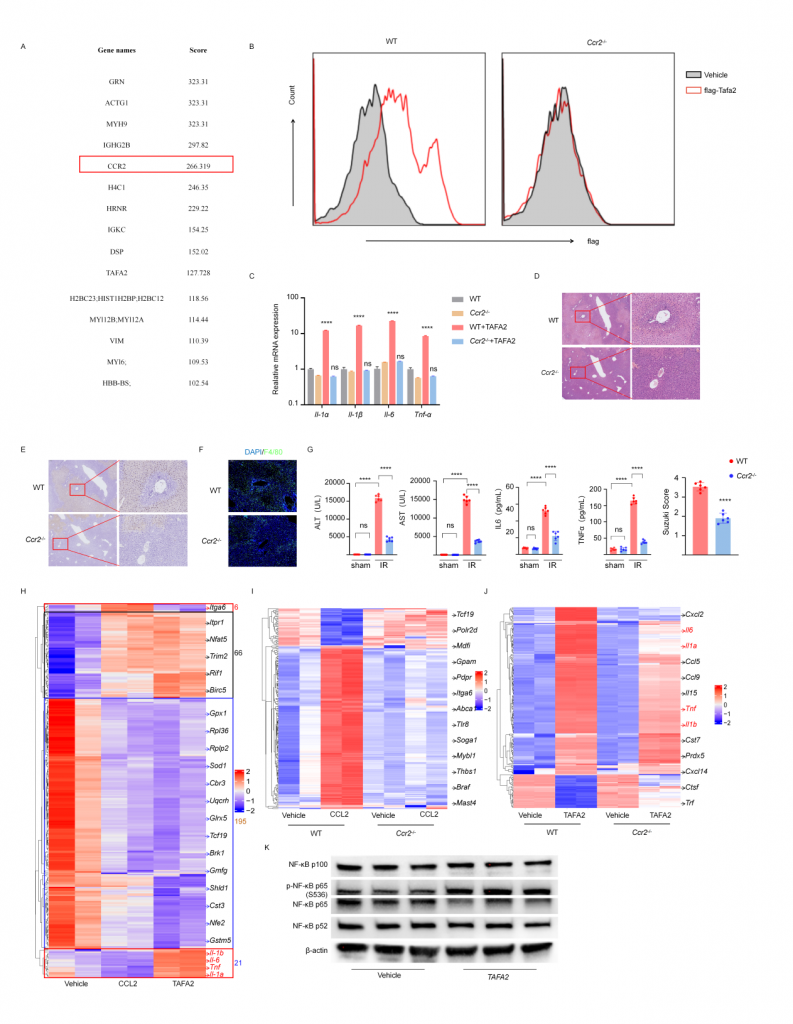
圖6-TAFA2與巨噬細(xì)胞表面CCR2互作
5.酸刺激減輕人類肝切除術(shù)中肝IRI
為了確認(rèn)酸刺激在人類肝IRI中的作用,研究者進行了開放、隨機、空白對照臨床試驗(ChiCTR2400088096),包含多種良性/惡性肝腫瘤、肝外傷、膿腫、囊腫和包蟲病,由于手術(shù)期間門靜脈短暫阻斷以減少肝切除過程中出血,導(dǎo)致肝出現(xiàn)缺血再灌注損傷。干預(yù)組33名患者,術(shù)前24h開始,每8h給與新鮮的25mM檸檬酸(持續(xù)5min),對照組33名患者不接受酸刺激,剔除6名術(shù)中肝缺血超過30min患者以及4名依從性差患者,肝切除術(shù)后第1/3/5天對患者肝功能進行評估,結(jié)果顯示酸刺激組血清ALT和AST水平顯著低于對照組,高ALT水平(>500 U/L)患者數(shù)量也顯著更低,這些結(jié)果表明酸刺激可減輕人類肝切除術(shù)引起的肝IRI。
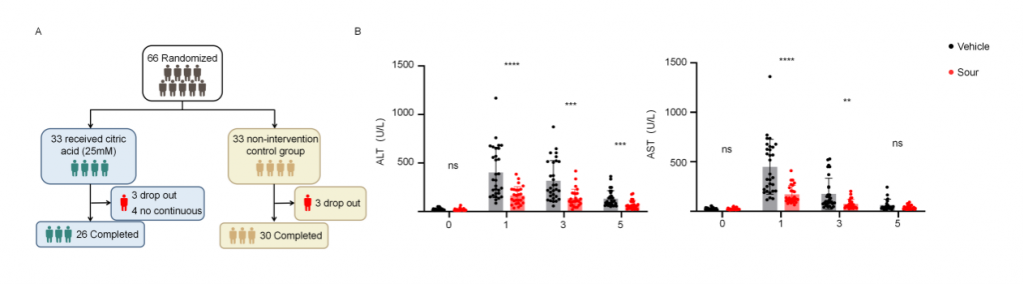
圖7-酸刺激減輕人類肝切除術(shù)中肝IRI
研究總結(jié)
該研究首次揭示了腦-肝軸在肝臟IRI中的調(diào)控作用,闡明了酸味刺激通過神經(jīng)信號通路緩解肝臟損傷的機制。研究不僅為肝臟IRI的治療提供了新的思路,還為神經(jīng)免疫調(diào)控在其他器官炎癥中的應(yīng)用奠定了基礎(chǔ)。研究團隊表示,未來將進一步探索神經(jīng)刺激療法在肝臟疾病中的應(yīng)用,特別是通過調(diào)控腦-肝軸來緩解肝臟炎癥和損傷。此外,研究團隊還將深入研究TAFA2蛋白的作用機制,開發(fā)針對TAFA2-CCR2信號通路的靶向藥物,為肝臟疾病的治療提供更多選擇。
]]>
豬不僅是重要的經(jīng)濟家畜,還在科研中扮演重要角色。豬器官與人類器官高度相似,被廣泛用于臨床前研究。
2020年,馬斯克展示了植入腦機芯片的豬,展示了腦電波信號的變化;2021年以來,全球已完成豬器官移植到腦死亡患者的實驗;2025年,空軍軍醫(yī)大學(xué)西京醫(yī)院又一次成功實施了豬肝臟異種移植到人腦死亡患者體內(nèi),與之前不一樣的是,這次是切除人類肝臟后只保留豬肝臟,探討豬肝能否完全替代人肝。
從時空實驗應(yīng)用條件看,豬的器官組織特征與人類接近,實驗開發(fā)難度較低。豬是二倍體生物,基因組質(zhì)量良好,有基因表達調(diào)控數(shù)據(jù)庫和單細(xì)胞數(shù)據(jù),便于跨物種比較研究。綜上,時空技術(shù)的應(yīng)用可提升研究精度,為后續(xù)研究提供精準(zhǔn)參考。
生殖方向
1.豬早期卵子發(fā)生的時空圖譜

英文標(biāo)題:Spatiotemporal dynamics of early oogenesis in pigs
發(fā)表期刊:Genome Biology
影響因子:10.1
發(fā)布時間:2025-01
DOI:10.1186/s13059-024-03464-8
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,豬E45、E55、E65、E75胚胎卵巢(n=2)。
空間轉(zhuǎn)錄組,豬E45、E55、E65、E75胚胎卵巢,10x Visium;E65胚胎卵巢,BMKMANU S1000。
百邁客生物為該研究提供了百創(chuàng)S1000空間轉(zhuǎn)錄組技術(shù)服務(wù)。
研究內(nèi)容總結(jié):
① 該研究結(jié)合單細(xì)胞RNA測序(scRNA-seq)和空間轉(zhuǎn)錄組學(xué)(ST)探索在豬卵子發(fā)生早期卵巢微環(huán)境的空間組織,構(gòu)建時空基因表達譜。將卵子發(fā)生不同階段的生殖細(xì)胞簇投射到空間圖譜中,揭示了發(fā)育中的豬卵巢中生殖細(xì)胞的“皮質(zhì)-髓質(zhì)(C-M)”分布。豬和人之間的跨物種分析揭示了在卵子發(fā)生過程中生殖細(xì)胞的保守的C-M分布模式,提示豬可以作為人類早期卵子發(fā)生過程的理想研究模型。
② 利用ST數(shù)據(jù)進行RNA速度分析,確定了豬卵巢皮質(zhì)和髓質(zhì)區(qū)顆粒細(xì)胞系的分子特征和空間動力學(xué)。通過空間共定位分析和細(xì)胞間通訊分析,揭示了皮質(zhì)和髓質(zhì)區(qū)域生殖細(xì)胞和體細(xì)胞之間獨特的細(xì)胞-細(xì)胞通訊模式。
③ 值得注意的是,卵巢組織的體外培養(yǎng)實驗結(jié)果證實細(xì)胞間NOTCH信號傳導(dǎo)和細(xì)胞外基質(zhì)(ECM)蛋白在啟動減數(shù)分裂和卵子形成程序中起關(guān)鍵作用,表明卵巢微環(huán)境對于生殖細(xì)胞的命運調(diào)控起著重要作用。

圖1-基于Cell2location,使用scRNA-seq數(shù)據(jù)對ST數(shù)據(jù)進行解卷積
2.時空技術(shù)解析豬滋養(yǎng)層類器官與母胎界面細(xì)胞多樣性

英文標(biāo)題:Defining Cellular Diversity at the Swine Maternal-Fetal Interface Using Spatial Transcriptomics and Organoids
發(fā)表期刊:bioRxiv
發(fā)布時間:2024-10
DOI:10.1101/2024.10.21.619461
實驗設(shè)計時空部分:單細(xì)胞轉(zhuǎn)錄組,滋養(yǎng)層細(xì)胞系類器官(n=3)。空間轉(zhuǎn)錄組:G65臍帶殘端附近1cm*1cm完整胎盤(包括母體和子代組織,n=4)。
研究內(nèi)容總結(jié):
① 研究者使用足月豬胎盤構(gòu)建的sTO(豬滋養(yǎng)層類器官)可以擴增、凍存并成功復(fù)蘇,高表達滋養(yǎng)層標(biāo)志基因KRT18、ELF3以及GATA3,與豬胎盤表達標(biāo)志更相似。此外,在人工基底膜中培養(yǎng)立刻形成懸浮培養(yǎng)可逆的局部頂面,通過和體內(nèi)較為一致的基因表達和細(xì)胞通訊程序分化成不同滋養(yǎng)層細(xì)胞。與當(dāng)前其他可用的體外模型相比,研究者開發(fā)的sTO是更理想的體外研究模型。
② 使用空間轉(zhuǎn)錄組技術(shù)定義妊娠中期豬母胎界面上滋養(yǎng)層原位轉(zhuǎn)錄組圖譜,無需依賴傳統(tǒng)標(biāo)志,揭示了豬子宮和胎盤的新marker,可用于精準(zhǔn)定義豬母胎界面組織學(xué)結(jié)構(gòu);數(shù)據(jù)還包含以往未分析到的腺窩區(qū)(areola)和交界區(qū),定義出3種腺窩細(xì)胞亞群,例如主要位于胎盤間質(zhì)附近而不明顯與交界區(qū)直接接觸的Areola-1。總之,該圖譜為后續(xù)豬生殖研究提供了基礎(chǔ)參考。
③ 空間數(shù)據(jù)可用于注釋sTO單細(xì)胞轉(zhuǎn)錄組數(shù)據(jù)中的細(xì)胞類型,進一步分析發(fā)現(xiàn)sTO涵蓋了豬胎盤中滋養(yǎng)層細(xì)胞群的發(fā)育和功能差異,sTO和胎盤組織具有相同保守的關(guān)鍵信號通路。此外,由于sTO中有更多增殖干細(xì)胞,因而僅在sTO單細(xì)胞數(shù)據(jù)中觀察到增殖干細(xì)胞與其他滋養(yǎng)層間存在互作。總之,這些凸顯了sTO在研究豬胎盤發(fā)育中的用途。

圖2-空間轉(zhuǎn)錄組學(xué)解析豬母胎界面的全局表達情況
3.豬竇卵泡的單細(xì)胞圖譜
 英文標(biāo)題:Deciphering Cellular Heterogeneity and Communication Patterns in Porcine Antral Follicles by Single-Cell RNA Sequencing
英文標(biāo)題:Deciphering Cellular Heterogeneity and Communication Patterns in Porcine Antral Follicles by Single-Cell RNA Sequencing
發(fā)表期刊:Animals
發(fā)布時間:2023-09
DOI:10.3390/ani13193019
實驗設(shè)計時空部分:單細(xì)胞轉(zhuǎn)錄組:D210母豬(取樣前48h內(nèi)注射5 IU PMSG)竇卵泡(n=2)。
研究內(nèi)容總結(jié):
① 單細(xì)胞數(shù)據(jù)揭示了豬竇卵泡內(nèi)細(xì)胞的顯著異質(zhì)性,特別是顆粒細(xì)胞,研究者首次在豬中確認(rèn)壁顆粒細(xì)胞(mGC)和卵丘細(xì)胞(nGC)存在不同亞群,例如mGC1亞群在激素信號轉(zhuǎn)導(dǎo)中起到重要作用,而mGC2主要負(fù)責(zé)雌激素合成,cGC2負(fù)責(zé)糖酵解和卵丘擴張,cGC1亞群具有mGC和cGC兩種顆粒細(xì)胞特征,表明卵泡內(nèi)存在多種狀態(tài)和功能的細(xì)胞。
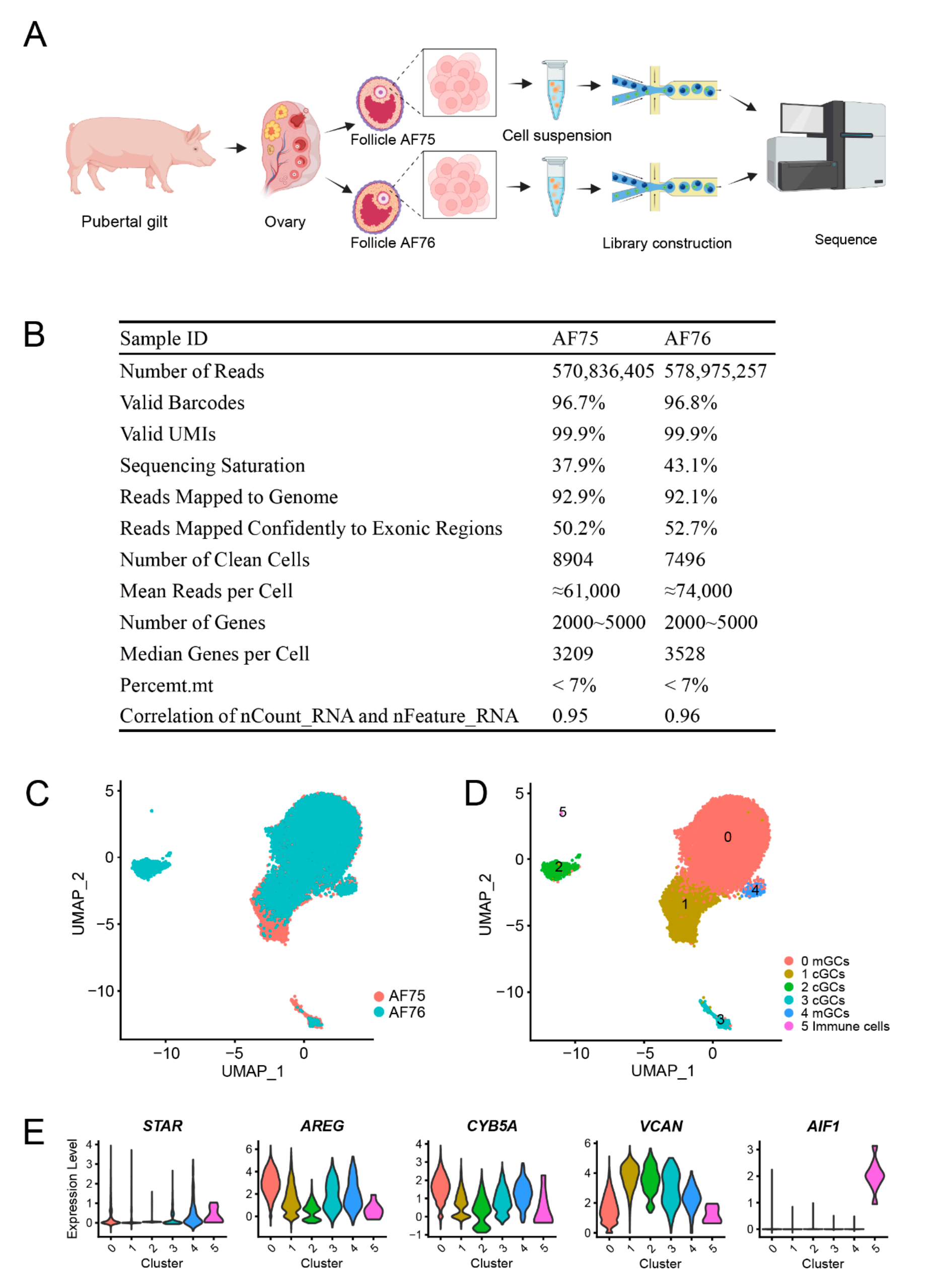
圖3-豬竇狀卵泡中細(xì)胞的降維聚類結(jié)果
發(fā)育方向
4.豬皮膚早期發(fā)育的時空圖譜

英文標(biāo)題:Integrating Single-Cell and Spatial Transcriptomics Reveals Heterogeneity of Early Pig Skin Development and aSubpopulation with Hair Placode Formation
發(fā)表期刊:Advanced Science
影響因子:14.3
發(fā)布時間:2024-04
DOI:10.1002/advs.202306703
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,E37胚胎(n=1,未知U),E41&E52&E85胚胎(每時期n=2,同窩有毛N、無毛H各1)。
空間轉(zhuǎn)錄組,E37胚胎(n=1),E41&E52&E85胚胎(每時期n=2,同窩有毛N、無毛H各1)。
研究內(nèi)容總結(jié):
① 單細(xì)胞數(shù)據(jù)分析得到7種主要細(xì)胞類型,空間轉(zhuǎn)錄組數(shù)據(jù)分析得到6種主要細(xì)胞類型,包括免疫細(xì)胞、內(nèi)皮細(xì)胞、周細(xì)胞、施旺細(xì)胞等,且不同類型細(xì)胞空間分布與已知的皮膚組織解剖結(jié)構(gòu)相一致,相關(guān)性分析表明單細(xì)胞數(shù)據(jù)和空間數(shù)據(jù)中鑒定出的細(xì)胞類型一致性較高。
② 聯(lián)合已發(fā)表的人、小鼠皮膚單細(xì)胞數(shù)據(jù)分析,發(fā)現(xiàn)豬表皮細(xì)胞、免疫細(xì)胞以及周細(xì)胞表達更多保守的特異性標(biāo)志基因,這些基因也與人類皮膚疾病相關(guān),表明與小鼠相比,豬皮膚或許是一種更合適的人皮膚疾病研究模型。
③ 表皮細(xì)胞簇進一步聚類得到9種亞型,OGN+/UCHL1+ 細(xì)胞主要在E37U中存在,毛囊間表皮(IFE)基底層細(xì)胞與祖細(xì)胞從E41到E52表現(xiàn)出增加趨勢。擬時序分析結(jié)果顯示存在兩種分化軌跡,OGN+/UCHL1+ 細(xì)胞—>IFE基底層細(xì)胞和真皮細(xì)胞前體(pre-DC)、OGN+/UCHL1+ 細(xì)胞—>成熟毛囊基底(Pc)發(fā)育和毛囊(HF)及角質(zhì)細(xì)胞分化。真皮成纖維細(xì)胞簇進一步聚類得到乳頭狀成纖維細(xì)胞和網(wǎng)狀成纖維細(xì)胞,進一步分析在E37U中鑒定出成纖維祖細(xì)胞。
④ 有毛豬中Pc的形成時期為E37-E41,無毛豬中缺乏Pc細(xì)胞。CytoTRACE、逆時序分析等結(jié)果表明,無毛豬OGN+/UCHL1+ 細(xì)胞增殖和遷移異常導(dǎo)致Pc不能正常形成,BMP和TGFβ是引起OGN+/UCHL1+ 細(xì)胞形成Pc的首個信號通路。

圖4-ST和scRNA-seq鑒定的常見細(xì)胞類型的轉(zhuǎn)錄組之間具有高度一致性
5.豬出生后肝臟發(fā)育的單細(xì)胞動態(tài)圖譜

英文標(biāo)題:Single-cell dynamics of liver development in postnatal pigs
發(fā)表期刊:Science Bulletin
影響因子:18.8
發(fā)布時間:2023-09
DOI:10.1016/j.scib.2023.09.021
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,出生后不同胎齡豬肝臟,D30(n = 3),D42 (n = 3),D150(n = 1),D730(n = 2)。
單細(xì)胞核ATAC,D240(n=1)。
研究內(nèi)容總結(jié):
① 構(gòu)建迄今最全面的出生后豬肝臟單細(xì)胞發(fā)育圖譜,涵蓋斷奶前(30天)、斷奶后(42天)、生長高峰(150天)和成年階段(730天)這四個生長發(fā)育重要階段,數(shù)據(jù)涵蓋單細(xì)胞轉(zhuǎn)錄組、單細(xì)胞核轉(zhuǎn)錄組以及單細(xì)胞ATAC數(shù)據(jù),共鑒定到23種細(xì)胞類型,包括肝臟中的三種稀有細(xì)胞類型,漿細(xì)胞樣樹突狀細(xì)胞(pDCs),CAVIN3+IGF2+內(nèi)皮細(xì)胞和EBF1+成纖維細(xì)胞。
② 發(fā)現(xiàn)斷奶前仔豬和成年豬脂肪酸合成的差異,擬時序分析鑒定出5693個基因在三個發(fā)育階段表現(xiàn)出顯著表達水平變化,進一步分析發(fā)現(xiàn)33個階段特異性轉(zhuǎn)錄因子,例如D30富集參與調(diào)節(jié)出生后肝細(xì)胞成熟的轉(zhuǎn)錄因子EZH2,D42富集參與調(diào)節(jié)晝夜節(jié)律的CLOCK,還首次鑒定出成年豬肝竇內(nèi)皮特異性的轉(zhuǎn)錄因子LUAP2。通路富集分析結(jié)果顯示不同發(fā)育階段肝竇內(nèi)皮細(xì)胞富集的通路不同,D40是免疫相關(guān)通路富集,D730中主要是代謝相關(guān)通路基因表達上調(diào)。
③ 豬D30免疫細(xì)胞主要是NK(自然殺傷細(xì)胞)和T細(xì)胞,以往有報道表明新生小鼠免疫系統(tǒng)重髓系細(xì)胞占比更大,出生D30豬肝臟免疫系統(tǒng)可能發(fā)生了從髓系細(xì)胞到淋巴系細(xì)胞的免疫系統(tǒng)轉(zhuǎn)變。此外,豬trNK(組織駐留自然殺傷細(xì)胞)具有與人類相似的轉(zhuǎn)錄因子表達特征,或表明豬可以作為研究人trNK的理想模型。

圖5-單細(xì)胞RNA測序(scRNA-seq)和單核RNA測序(snRNA-seq)鑒定發(fā)育中肝臟細(xì)胞類型
免疫方向
6.豬腎異種移植排異的時空圖譜

英文標(biāo)題:Spatiotemporal immune atlas of a clinicalgrade gene-edited pig-to-human kidney xenotransplant
發(fā)表期刊:Nature Communications
影響因子:14.7
發(fā)布時間:2024-04
DOI:10.1038/s41467-024-47454-7
實驗設(shè)計時空部分:
單細(xì)胞核轉(zhuǎn)錄組,移植前豬腎穿刺組織,移植后24h/72h/74h豬腎穿刺組織(n=4)。單細(xì)胞轉(zhuǎn)錄組,移植74h后切除的腎組織CD45+細(xì)胞(n=1);
空間轉(zhuǎn)錄組,移植前豬腎穿刺組織,移植后24h/72h/74h豬腎穿刺組織(n=4)。
研究內(nèi)容總結(jié):
① 基因編輯豬腎移植給腦死亡且雙腎切除患者。數(shù)據(jù)分析結(jié)果顯示,未在異種移植后的豬腎組織中發(fā)現(xiàn)急性細(xì)胞排異或IgM/IgG/配體蛋白結(jié)合的證據(jù),但發(fā)現(xiàn)急性腎小管壞死和病因不明的血栓性微血管病變。
② 移植后豬腎中髓系細(xì)胞是人和豬腎中檢測到的最多的免疫細(xì)胞,移植3天內(nèi)人B細(xì)胞和T淋巴細(xì)胞很少檢測到,3天后檢測到的人免疫細(xì)胞增多但豐度遠(yuǎn)小于豬免疫細(xì)胞。
③ 移植3天后發(fā)現(xiàn)人類中性粒細(xì)胞和單核細(xì)胞主要與豬內(nèi)皮細(xì)胞共定位,人巨噬細(xì)胞主要與豬基質(zhì)細(xì)胞共定位,表明人免疫細(xì)胞對豬腎皮質(zhì)浸潤有限。
④ 移植豬腎中人和豬巨噬細(xì)胞更傾向于激活抗炎癥基因表達。
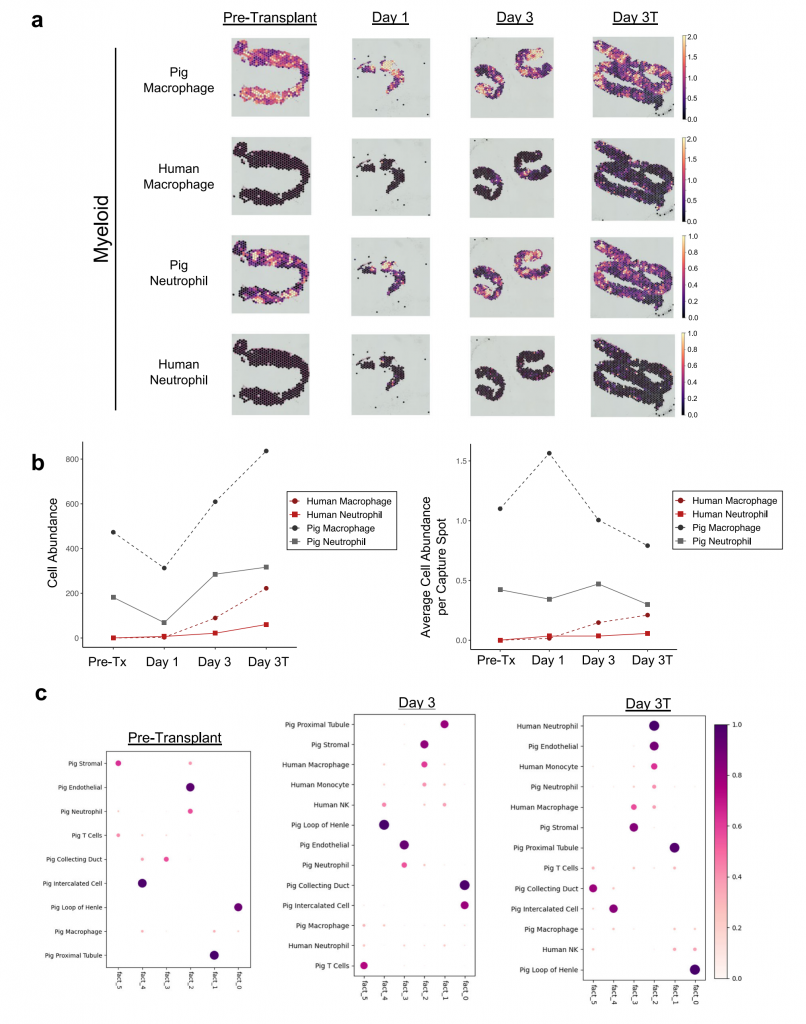
圖6-人髓系細(xì)胞對豬腎異種移植的有限浸潤
7.豬空腸免疫細(xì)胞單細(xì)胞圖譜
?
英文標(biāo)題:Single-Cell Transcriptional Analysis of Lamina Propria Lymphocytes in the Jejunum Reveals Innate Lymphoid Celllike Cells in Pigs
發(fā)表期刊:The Journal of Immunology
發(fā)布時間:2024-01
DOI:10.4049/jimmunol.2300463
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,4周大仔豬,空腸固有層淋巴細(xì)胞(n=3),空腸先天淋巴樣細(xì)胞(n=3)。
研究內(nèi)容總結(jié):
② 豬空腸ILC標(biāo)志基因與人基因相似,包括IL-23R、AHR、NCR2等。此外,研究者發(fā)現(xiàn)ILC3可能分化成ILC2、ILC1和NK細(xì)胞。ILC3亞群種,ILC3b沒有明顯高表達基因,是ILC亞群中具有更高轉(zhuǎn)分化可塑性的亞群,IKZF1和TGFB1可能與該過程有關(guān)。
③ 豬ILC3表達TAC3基因,該基因編碼蛋白的受體是NK3R,可以促進釋放促性腺激素釋放激素,繼而促進性激素釋放,因此TAC3在促性腺激素軸上起到重要作用,因而可能存在腸/性腺軸,暗示豬腸道ILC3或調(diào)控性腺發(fā)育。

圖7-豬空腸中l(wèi)LC3s的分型研究
8.感染PEDV仔豬的空腸單細(xì)胞圖譜
?
英文標(biāo)題:Identification of Cell Types and Transcriptome Landscapes of Porcine Epidemic Diarrhea VirusInfected Porcine Small Intestine Using Single-Cell RNA Sequencing
發(fā)表期刊:The Journal of Immunology
發(fā)布時間:2023-02
DOI:10.4049/jimmunol.2101216
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,口服2ml PEDV-24h的3天大仔豬(n=1,4只豬樣本混合),口服對照液體-24h的3天大仔豬(n=1,4只豬樣本混合)
研究內(nèi)容總結(jié):
① 首次繪制感染PEDV(豬流行性腹瀉病毒)的仔豬小腸單細(xì)胞圖譜,使用人小腸細(xì)胞類型marker鑒定出12種細(xì)胞類型,發(fā)現(xiàn)豬小腸tuft細(xì)胞新特異性標(biāo)志基因DNAH11,結(jié)果表明多數(shù)人小腸特異性marker也可以用到豬研究中。此外,豬小腸Th17細(xì)胞(3和9簇)特異性高表達IL17A、IL7F和IL22,但不高表達T細(xì)胞代表性CD3,這可能與豬T細(xì)胞分化有關(guān)。
② 抗菌肽(AMP)相關(guān)基因分析結(jié)果顯示,僅DEFB115和REG3G在仔豬空腸部分的腸細(xì)胞中最為豐富,PEDV感染會導(dǎo)致REG3G顯著上調(diào)。REG3G表達與IL33、MyD88、STAT3等相關(guān),在體外實驗中發(fā)現(xiàn)感染PEDV后IPEC-J2細(xì)胞IL33表達水平以及STAT3磷酸化水平顯著增加,表明IL33-STAT3信號通路可能在PEDV感染誘導(dǎo)的REG2G表達中起到重要作用。
③ 功能富集分析結(jié)果顯示,病毒感染導(dǎo)杯狀細(xì)胞、tuft細(xì)胞和腸內(nèi)分泌細(xì)胞中緊密連接和粘附連接通路水平顯著降低,體外實驗確認(rèn)PEDV感染的IPEC-J2細(xì)胞中,緊密連接通路相關(guān)基因表達顯著降低,但感染晚期粘附連接通路相關(guān)基因轉(zhuǎn)錄和蛋白質(zhì)水平表達顯著增加,這可能是由于不同腸細(xì)胞類型對PEDV感染的不同響應(yīng)導(dǎo)致的。冠狀病毒受體分析結(jié)果表明,豬小腸上皮細(xì)胞高表達多種不同的冠狀面病毒受體,這一發(fā)現(xiàn)支持豬易受冠狀病毒感染并表現(xiàn)感染相關(guān)腸道癥狀。

圖8-豬小腸空腸段細(xì)胞類型的測定
同源器官比較
9.脊椎動物肺單細(xì)胞圖譜

英文標(biāo)題:Origin and stepwise evolution of vertebrate lungs
發(fā)表期刊:Nature ecology & evolution
影響因子:14.1
發(fā)布時間:2025-2
DOI:10.1038/s41559-025-02642-6
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組測序:
塞內(nèi)加爾多鰭魚,肺(n=2),腦/食管/鰓/心臟/腸/肌肉/胃(各器官,n=1);
非洲肺魚,肺(n=2),腦/食管/鰓/心臟/腸/腎/肌肉/皮膚/胃(各器官,n=1);
條紋斑竹鯊,壺腹/魚鰭/心臟/腸/胰腺/鰓/牙(各器官,n=3),腦/食管/腎(各器官,n=2),脾臟(n=6),眼睛(n=1);雞肺,E6/E7/P3(各時期,n=1);
非洲牛蛙肺(n=2);鬃獅蜥肺(n=1)。
人/小鼠/大鼠/豬的肺數(shù)據(jù)使用已有的單細(xì)胞轉(zhuǎn)錄組測序數(shù)據(jù)。
百邁客生物為塞內(nèi)加爾多鰭魚和非洲肺魚除肺以外的組織,提供了百創(chuàng)DG1000單細(xì)胞轉(zhuǎn)錄組測序服務(wù)。
研究內(nèi)容總結(jié):
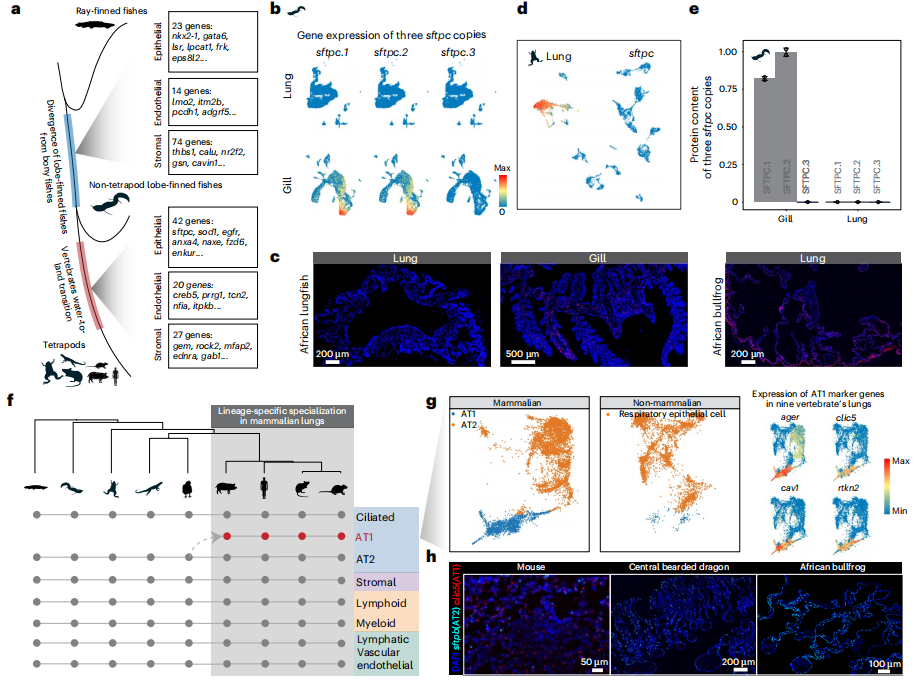
① 使用單細(xì)胞轉(zhuǎn)錄組測序分析9種動物的成體/胚胎肺組織及其他組織,發(fā)現(xiàn)軟骨魚具有多個肺發(fā)育所必須得遺傳組分,包括肺特異性基因,但不同物種中這些基因的表達模式和功能可能有差異,表明軟骨魚距離擁有肺在“進化上只差一步之遙”;軟骨魚食管和胃中,少量細(xì)胞共表達對呼吸脊椎動物肺功能十分重要的sftpb和abca3基因,暗示頜類脊椎動物最近共同祖先(LCA)經(jīng)歷顯著的演化變化,不僅發(fā)育出重要特征如頜和成對附件,還為肺最終出現(xiàn)奠定基礎(chǔ)。
② 許多保守的非編碼組分(CNE)來源于頜類脊椎動物祖先,暗示肺的遺傳基礎(chǔ)可能在脊椎動物演化很早就出現(xiàn),但無肺蠑螈中缺失的CNE與其促肺活性無關(guān),表明這些組分可能還具有除肺發(fā)育以外的多種功能;硬骨魚祖先來源的CNE在無肺蠑螈表現(xiàn)出更高的丟失率,也反映出這些CNE具有顯著的肺功能特異性,強調(diào)了復(fù)雜器官起源中調(diào)控組分演化的重要性。這些證據(jù)暗示肺起源的兩個階段過程,最開始頜類脊椎動物L(fēng)CA出現(xiàn)基礎(chǔ)的肺相關(guān)遺傳組分,隨后譜系演化出更特異性的肺增強子產(chǎn)生硬骨魚;功能完全的肺可能在軟骨魚和硬骨魚譜系分開后演化而來。
③ 全基因組復(fù)制對肺演化很重要,1866個肺相關(guān)直系同源基因中有776個是脊椎動物兩輪全基因組復(fù)制(2R-WGD)的產(chǎn)物;哺乳動物譜系特異的基因復(fù)制也十分重要,sfta2-/-小鼠表現(xiàn)出明顯的呼吸疾病,包括顯著的炎癥。
④ 研究成果支持了兩個關(guān)于復(fù)雜器官的前期假設(shè)。首先,肺進化是一個逐步過程,與達爾文預(yù)測相一致,與眼睛和其他器官類似,肺演化似乎是隨時間逐漸發(fā)生的。其次,反映了Jacob定律,即演化類似“修補匠”,使用已有的遺傳基礎(chǔ),包括招募、征用已經(jīng)存在的基因和調(diào)控組分。總之,這些結(jié)果表明調(diào)控網(wǎng)絡(luò)的修改對肺的起源和演化是十分重要的,新基因或基因重復(fù)的出現(xiàn)提供了基礎(chǔ)材料,即使這些基因并不會立刻發(fā)揮作用。
10.野豬/萊蕪豬/杜洛克豬新生骨骼肌單細(xì)胞圖譜
 英文標(biāo)題:Single-Cell RNA-Sequencing Provides Insight into Skeletal Muscle Evolution during the Selection of Muscle Characteristics
英文標(biāo)題:Single-Cell RNA-Sequencing Provides Insight into Skeletal Muscle Evolution during the Selection of Muscle Characteristics
發(fā)表期刊:Advanced Science
影響因子:14.3
發(fā)布時間:2023-10
研究內(nèi)容總結(jié):
① 使用單細(xì)胞轉(zhuǎn)錄組測序技術(shù)繪制了野豬、萊蕪豬和杜洛克豬新生骨骼肌駐留細(xì)胞圖譜,鑒定出9種細(xì)胞類型,包括成肌細(xì)胞、肌細(xì)胞、衛(wèi)星細(xì)胞等,定義兩種新亞型,MT豐富FAP(?bro-adipogenic progenitors)以及肌細(xì)胞樣FAP。
② 與不同物種的胎兒和成體骨骼肌相比,豬新生骨骼肌細(xì)胞組成類型更豐富。例如,豬新生骨骼肌衛(wèi)星細(xì)胞不僅包含衛(wèi)星干細(xì)胞(PAX7+),還包括兩個有不同肌源性潛能的亞群(HES1+和TRIB1+衛(wèi)星細(xì)胞);少有研究可解答骨骼肌中FAP的異質(zhì)性,但本研究發(fā)現(xiàn)4種FAP亞群。與人、小鼠骨骼肌數(shù)據(jù)聯(lián)合分析,發(fā)現(xiàn)豬骨骼肌駐留細(xì)胞的通用marker和物種特異性的marker。
③ 豬新生骨骼肌中發(fā)現(xiàn)增殖分化活性狀態(tài)的干性樣細(xì)胞,如pro-NK/T、間充質(zhì)干細(xì)胞(MSC)、間質(zhì)細(xì)胞等,且MSC和FAP間存在一群具有典型標(biāo)志基因如CD73、CD90和PDGFRA的連續(xù)態(tài)細(xì)胞亞群。此外,擬時序分析揭示杜洛克豬新生骨骼肌中的成肌譜系干細(xì)胞處于初始階段,野豬的成肌譜系已處于分化末期和成熟期,萊蕪豬位于中間態(tài)。
④ 不同品種豬具有不同骨骼肌表型,它們骨骼肌駐留細(xì)胞譜系存在一定差異。與家豬相比,野豬缺少兩種FAP亞群,但存在THY1+衛(wèi)星細(xì)胞;發(fā)現(xiàn)品種特異性細(xì)胞類型,例如萊蕪豬具有COL13A1+ 腱細(xì)胞;與萊蕪豬和野豬相比,杜洛克豬新生肌肉中具有更多增殖的前體脂肪細(xì)胞,或意味杜洛克豬未來選育后可積累更高的IMF(肌內(nèi)脂肪)。
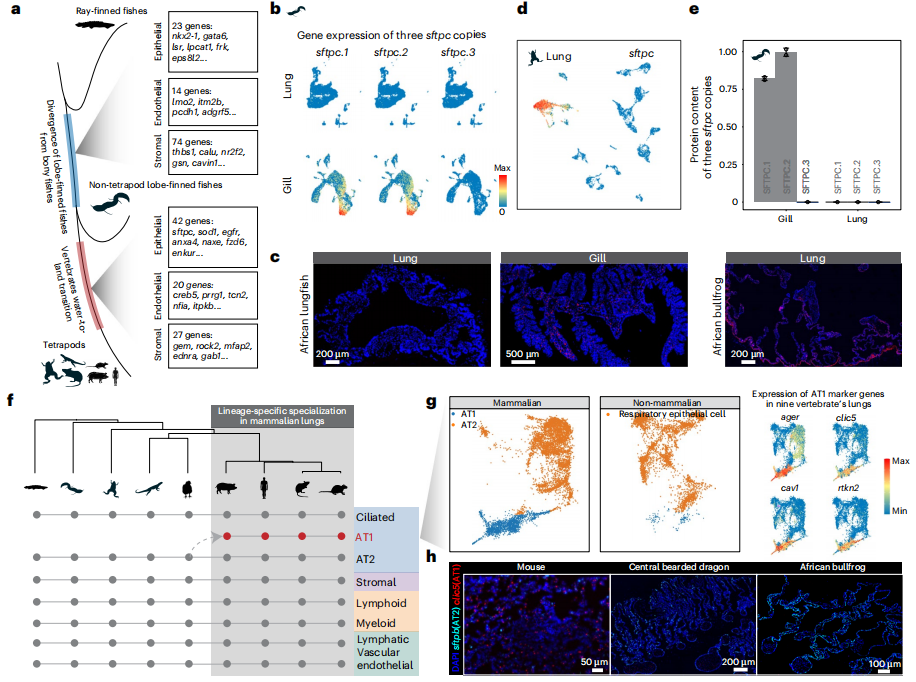
圖9-野豬、萊蕪豬和杜洛克豬骨骼肌細(xì)胞的單細(xì)胞轉(zhuǎn)錄譜分析
11.人/小鼠/大鼠/豬胃竇跨物種比較單細(xì)胞圖譜

英文標(biāo)題:Cross-species single-cell transcriptomic analysis of animal gastric antrum reveals intense porcine mucosal immunity
發(fā)表期刊:Cell Regeneration
發(fā)布時間:2023-08
DOI:10.1186/s13619-023-00171-w
實驗設(shè)計時空部分:單細(xì)胞轉(zhuǎn)錄組,人(n=4),小鼠(n=3),大鼠(n=3),豬(n=3)。
研究內(nèi)容總結(jié):
① 使用單細(xì)胞轉(zhuǎn)錄組測序技術(shù)構(gòu)建人、豬、大鼠和小鼠的胃竇單細(xì)胞轉(zhuǎn)錄組圖譜,鑒定出9種類型的細(xì)胞,包括小凹黏膜細(xì)胞、小凹祖細(xì)胞、基底腺粘液細(xì)胞等。絕大多數(shù)類型的細(xì)胞存在于兩個或者多個物種中,F(xiàn)3和CLCA1低表達水平的祖細(xì)胞僅在人類和豬的胃竇上皮中發(fā)現(xiàn),高表達F3的基底粘液腺細(xì)胞僅在豬胃竇上皮中發(fā)現(xiàn),通路富集分析表明F3+細(xì)胞類群可能與細(xì)胞遷移、細(xì)胞增殖以及蛋白穩(wěn)定維持相關(guān)。
② 功能富集分析結(jié)果表明,人的胃竇上皮高表達金屬離子穩(wěn)態(tài)相關(guān)基因,豬的胃竇上皮高表達免疫相關(guān)基因,大鼠和小鼠的胃竇上皮高表達脂代謝相關(guān)基因,這種差異可能是由于飲食習(xí)慣導(dǎo)致的。豬胃竇上皮類器官bulk RNA-seq數(shù)據(jù)也確認(rèn)這一發(fā)現(xiàn),進一步的豬胃竇上皮細(xì)胞類器官體外實驗結(jié)果表明,TNFα能夠特異性地上調(diào)T細(xì)胞和B細(xì)胞活化相關(guān)基因,這些結(jié)果表明,正常生理狀態(tài)下豬胃竇上皮細(xì)胞具有強免疫能力,可能與其復(fù)雜飲食習(xí)慣和居住環(huán)境有關(guān)。
③ 進一步分析人和豬胃竇中的免疫細(xì)胞,發(fā)現(xiàn)人胃竇免疫細(xì)胞高表達B細(xì)胞/T細(xì)胞激活和功能相關(guān)基因,而豬胃竇顯著高表達B細(xì)胞/T細(xì)胞細(xì)胞增殖相關(guān)基因;細(xì)胞通訊分析結(jié)果顯示,人胃竇中上皮細(xì)胞和基質(zhì)細(xì)胞間高表達或特異性表達免疫細(xì)胞產(chǎn)生、成熟、維持和功能相關(guān)受配體對,而豬中主要是與上皮細(xì)胞生長分化、抗炎癥和抗菌、免疫細(xì)胞增殖相關(guān)。

圖10-scRNA-seq分析揭示了人、豬大鼠和小鼠胃竇細(xì)胞組成及基因表達譜
基礎(chǔ)圖譜
12.豬器官全景單細(xì)胞圖譜

英文標(biāo)題:Endothelial cell heterogeneity and microglia regulons revealed by a pig cell landscape at single-cell level
發(fā)表期刊:Nature Communications
影響因子:14.7
發(fā)布時間:2022-06
DOI:10.1038/s41467-022-31388-z
實驗設(shè)計時空部分:
單細(xì)胞轉(zhuǎn)錄組,6個月大豬,肝(n=1),血液PBMC(n=1),視網(wǎng)膜(n=1),脾臟(n=1),腸道(n=1),肺(n=1),脂肪組織(n=2)。
單細(xì)胞核轉(zhuǎn)錄組,6個月大豬,腦(n=9),視網(wǎng)膜(n=1),腎臟(n=1),心臟(n=1),脾臟(n=1),肝臟(n=1),肺(n=1)。
研究內(nèi)容總結(jié):
① 使用單細(xì)胞轉(zhuǎn)錄組測序及單細(xì)胞核轉(zhuǎn)錄組測序,構(gòu)建首個家豬多器官單細(xì)胞圖譜,得到234種降維聚類簇,鑒定出58種細(xì)胞類型及其相關(guān)的顯著富集標(biāo)志基因,搭建了可視化家豬單細(xì)胞轉(zhuǎn)錄組數(shù)據(jù)庫(Pig Single Cell Atlas Database)。
② 對血管內(nèi)皮細(xì)胞數(shù)據(jù)進一步分析,鑒定出21種具有特異表達特征和功能的血管內(nèi)皮細(xì)胞類型,包括脂肪組織中依賴于TGF-b2信號通路內(nèi)皮-間充質(zhì)轉(zhuǎn)化亞型;人/豬肝臟、腎臟和心臟組織中,內(nèi)皮細(xì)胞主要通過VEGF、PDGF,TGF-β和BMP通路與其他細(xì)胞類型互作,但不同細(xì)胞類型的互作通路有差異,例如豬內(nèi)皮細(xì)胞可與肝臟免疫細(xì)胞、心臟所有細(xì)胞細(xì)胞類型通過PDGF交流,但在腎臟中可通過PDGF通路與內(nèi)皮細(xì)胞交流的僅有足細(xì)胞。總之,這些結(jié)果表明家豬單細(xì)胞圖譜為研究豬或人組織中細(xì)胞間互作提供了有參考價值的基礎(chǔ)數(shù)據(jù)。
③ 對家豬、人、小鼠、猴、倉鼠、栗鼠、鼴鼠等13個物種的大腦小膠質(zhì)細(xì)胞單細(xì)胞轉(zhuǎn)錄組數(shù)據(jù)進行分析,發(fā)現(xiàn)1590個保守的轉(zhuǎn)錄組因子(TF)靶向?qū)Γ渲蠱EF2C、SPI1、IRF8、ZFP36L1在13個物種的小膠質(zhì)細(xì)胞中均高表達,這些結(jié)果表明家豬單細(xì)胞圖譜為研究不同物種小膠質(zhì)細(xì)胞保守和差異的遺傳調(diào)控網(wǎng)絡(luò)提供了基礎(chǔ)數(shù)據(jù),對未來腦小膠質(zhì)細(xì)胞功能研究發(fā)展具有重要意義。

圖11-豬20種組織單細(xì)胞圖譜
從以上文章可以看到,豬相關(guān)研究除了養(yǎng)殖、繁育、品種優(yōu)化、營養(yǎng)學(xué)、表型性狀、遺傳演化等方面,生殖、發(fā)育、衰老、損傷與修復(fù)、異種移植、神經(jīng)學(xué)等方向研究成果也具有較高參考和轉(zhuǎn)化價值。未來,可能會出現(xiàn)豬睪丸、胚胎發(fā)育、心血管組織、腦組織、疾病模型、異種移植/同種異體移植、鼻嗅覺、舌味覺、免疫系統(tǒng)等方向的研究成果。
Ps:關(guān)注時空組學(xué)在豬領(lǐng)域研究請聯(lián)系當(dāng)?shù)貥I(yè)務(wù)經(jīng)理獲取原文~
1. Ge W, Niu YL, et al. Spatiotemporal dynamics of early oogenesis in pigs. Genome Biol. 2025 Jan 2;26(1):2.
2. Cole R. McCutcheon, et al. Defining Cellular Diversity at the Swine Maternal-Fetal Interface Using Spatial Transcriptomics and Organoids. bioRxiv. 2024.
3. Chen N, et al. Deciphering Cellular Heterogeneity and Communication Patterns in Porcine Antral Follicles by Single-Cell RNA Sequencing. Animals (Basel). 2023 Sep 26;13(19):3019.
4. Wang Y, et al. Integrating Single-Cell and Spatial Transcriptomics Reveals Heterogeneity of Early Pig Skin Development and a Subpopulation with Hair Placode Formation. Adv Sci (Weinh). 2024 May;11(20):e2306703.
5. Rao L, et al. Single-cell dynamics of liver development in postnatal pigs. Sci Bull (Beijing). 2023 Nov 15;68(21):2583-2597.
6. Cheung MD, et al. Spatiotemporal immune atlas of a clinical-grade gene-edited pig-to-human kidney xenotransplant. Nat Commun. 2024 Apr 11;15(1):3140.
7. Wang J, et al. Single-Cell Transcriptional Analysis of Lamina Propria Lymphocytes in the Jejunum Reveals Innate Lymphoid Cell-like Cells in Pigs. J Immunol. 2024 Jan 1;212(1):130-142.
8. Fan B, et al. Identification of Cell Types and Transcriptome Landscapes of Porcine Epidemic Diarrhea Virus-Infected Porcine Small Intestine Using Single-Cell RNA Sequencing. J Immunol. 2023 Feb 1;210(3):271-282.
9. Li, Y., et al. Origin and stepwise evolution of vertebrate lungs. Nat Ecol Evol (2025).
10. Xu D, et al. Single-Cell RNA-Sequencing Provides Insight into Skeletal Muscle Evolution during the Selection of Muscle Characteristics. Adv Sci (Weinh). 2023 Dec;10(35):e2305080.
11. Wang X, et al. Cross-species single-cell transcriptomic analysis of animal gastric antrum reveals intense porcine mucosal immunity. Cell Regen. 2023 Aug 1;12(1):27.
12. Wang F, et al. Endothelial cell heterogeneity and microglia regulons revealed by a pig cell landscape at single-cell level. Nat Commun. 2022 Jun 24;13(1):3620.
]]>